
Profiling Genetic Breeding Progress in Bagrid Catfishes
by
 ,
,
Lishi Huang
1,†,
Guosong Zhang
2,†,
Yupeng Zhang
1,
Xinlan Li
1,
Zhong Luo
1,
Wenyu Liu
1,
Fu Luo
1,
Haifeng Liu
1,
Shaowu Yin
3,
Jun Jiang
1,
Xia Liang
2,* and
Quanquan Cao
1,* 1
College of Animal Science and Technology, Sichuan Agricultural University, Chengdu 611130, China
2
Key Laboratory for Physiology Biochemistry and Application, Heze University, Heze 274015, China
3
College of Marine Science and Engineering, College of Life Sciences, Nanjing Normal University, Nanjing 210023, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Fishes 2023, 8(8), 426; https://doi.org/10.3390/fishes8080426
Submission received: 25 June 2023
/
Revised: 3 August 2023
/
Accepted: 17 August 2023
/
Published: 21 August 2023
(This article belongs to the Special Issue Application and Perspectives in Fish Genetics Breeding)
Abstract
:The Bagridae fish family holds significant economic value and enjoys a high demand in the commercial market due to its desirable characteristics, such as delicious meat, fewer muscle spines, and a pleasing flavor. In recent years, the Bagridae has experienced rapid development in the seed industry in response to industry demands. Researchers have conducted extensive studies on genetic breeding in the Bagridae, employing a comprehensive breeding approach that combines conventional breeding techniques with innovative new technologies and cooperative efforts. In this review, we present an overview of the market situation and discuss various breeding processes applied to the Bagridae, including selective breeding, genetic engineering breeding, cell engineering breeding, molecular marker-assisted breeding, and hybrid breeding. Additionally, we focus on introducing the cultivation methods for two new Bagridae varieties. Furthermore, we explore and summarize the breeding conditions, genetic background, heterosis, comprehensive breeding technology, stress resistance, and metabolic nutrition of hybrid Bagridae catfishes. Overall, noticeable heterosis has been observed in the breeding of the hybrid yellow catfish “Huangyou-1”, which exhibits promising market prospects and economic benefits for aquaculture. Our study aims to underscore the importance of harnessing the heterosis of catfish, particularly the Pelteobagrus and Bagridae species. Consequently, this review provides an update on the current available information regarding the seed industry of the Bagridae fish family.
Key Contribution: Various breeding processes applied to the Bagridae were discussed including selective breeding, genetic engineering breeding, cell engineering breeding, molecular marker-assisted breeding, and hybrid breeding. And this review highlights progress in the research on the breeding conditions, genetic background, stress resistance, metabolic nutrition, and heterosis of hybrid Bagrid catfish.
1. Background
The Bagrid catfishes (family Bagridae) are freshwater fish comprising 245 species found in Africa and Asia [1]. According to the search volume on the websites Fishbase (https://www.fishbase.se/search.php, accessed on 15 June 2020) and FunDiving, the most typical feature of Bagrid catfishes is their possession of four pairs of well-developed barbels covered by a layer of taste bud-enriched epithelium [2]. Bagrid catfishes lack scales, and they have serrated pectoral fin spines and a dorsal fin preceded by a spine. Some species have a relatively long base on their adipose fin [1]. Larger Bagrids are important as food fish, and certain species are kept as aquarium fish [1], such as Pelteobagrus fulvidraco (Figure 1A), Pseudobagrus ussuriensis (Figure 1B), Leiocassis longirostris (Figure 1C), Pelteobagrus vachelli (Figure 1D), Tachysurus virgatus (Figure 1E), and Hyalobagrus ornatus (Figure 1F).
Within the Bagrid catfishes, species belonging to the genus Pelteobagrus, including P. vachelli, P. eupogon, and P. fulvidraco, are distributed in rivers and lakes of East and Southeast Asia, and some of them hold commercial importance [3,4]. Yellow catfish (Pelteobagrus fulvidraco), an indigenous aquaculture species in the Pelteobagrus genus, possesses characteristics such as adaptability, omnivorous diet, and high nutritional value. For instance, yellow catfish can survive in water temperatures ranging from 1 to 38 °C, and they are widely distributed in China, including the Yangtze River, Yellow River, Pearl River, and Heilongjiang River, except for the southwest and northwest plateau areas. Their diet consists of bottom-dwelling insects, mollusks, and occasionally fish [5]. Moreover, the production of yellow catfish has increased due to the availability of hatchery-produced seed, a standardized seed transportation system, and suitable feed [3]. They also possess desirable characteristics such as delicious meat, fewer muscle spines, and an appealing flavor. These attributes have contributed to yellow catfish’s popularity as a commercial fish [5,6,7].
In recent years, research on the genus Pelteobagrus has gained significant attention. According to analyses of the Web of Science, Pelteobagrus is valued in research on fisheries (28%), marine freshwater biology (19%), and biochemistry molecular biology (10%) (Figure 2A). The number of cited papers in this field has shown a rapid increase from 2011 to 2020 (Figure 2B). Specifically, according to the analysis of the Carrot database, which is a memory data type store, more studies are focusing on genetic studies, especially related to hybrids, genes, and mitochondrial genomes (Figure 2C).
China has also cultivated two new aquatic varieties via the Ministry of Agriculture, based on Pelteobagrus fulvidraco—hybrid “yellow catfish Huangyou No. 1” of P. fulvidraco ♀ × P. vachelli ♂ and “yellow catfish P. fulvidraco all-male No. 1”.
This review introduces the development of the Bagridae family and discusses the combination of various breeding methods, including selective breeding, gene engineering breeding, cell engineering breeding, and crossbreeding. The research progress of hybrid Bagrid catfishes is also presented, providing comprehensive knowledge for improving fish quality and yield. This contributes to alleviating the phenomenon of resource shortage and environmental destruction, promoting the development of sustainable and environmentally friendly aquaculture.
2. Development Status of the Aquaculture Industry for Yellow Catfish
Yellow catfish are cultured in various modes, primarily in ponds and cages [8]. Additionally, increasing production can be achieved through multi-age stocking, mixed-culture, and mixed breed rearing under monoculture systems [9]. Currently, pond monoculture has become the predominant approach [10]. However, the yellow catfish industry has been severely affected by several diseases. For instance, pathogen infections, enteric septicemia, ulcerative syndrome, ascites disease, red-head disease, and crack-head disease are among the diseases that can easily affect artificial aquaculture and lead to immune disturbances in yellow catfish [11]. High-density farming can increase the frequency of bacterial infections, including crack-head disease. Crack-head disease, a typical yellow catfish bacterial disease, can be caused by Gram-negative Edwardsiella. The dietary supplementation of vitamin D3 can reduce the death rate of yellow catfish after an Edwardsiella attack [12,13]. Moreover, excessive hepatic lipid deposition in yellow catfish has been widely reported to decrease disease resistance and harvest quality in many farms [14]. Therefore, it is essential to explore appropriate breeding methods based on the local ecological environment, reasonable bait supply, and disease prevention and control.
Over the past twenty years, freshwater aquaculture has become a pillar industry in mainland China’s agriculture, ranking first in the production and consumption of farmed freshwater species [9,15]. According to the China Fishery Statistical Yearbook 2020, yellow catfish production in several provinces of China is shown in Figure 3. Hubei Province leads in national yellow catfish production with 140,786 tons, followed by Zhejiang Province with 122,020 tons. Nine provinces have achieved production levels of more than 10,000 tons. In recent years, intensive high-density farming has gradually been adopted in yellow catfish farming, leading to higher economic benefits. For example, fishermen in Guangdong Province (the tropical region of China) make more money than those in Zhejiang Province in fishponds of the same size, benefiting from high-density aquaculture and an extended breeding season from May to September [6,9].
According to data from the Ministry of Agriculture of the People’s Republic of China (China Fishery Statistical Yearbook, http://english.moa.gov.cn/ accessed on 15 June 2020), the population of yellow catfish has tripled in a span of ten years from 2010 to 2019, specifically in terms of the demand for yellow catfish fry nationwide (7371 billion to 21,479 billion fish fries) (Figure 4A). Furthermore, thanks to advancements in breeding technology, the total production of yellow catfish in China reached a staggering 537 thousand tons in 2019, marking a fourfold increase from 2008 (134,448 tons) to 2019 (536,964 tons) (Figure 4B). The rise in production signifies an increasing demand for yellow catfish, but it does not necessarily imply that the current fry quality of yellow catfish is satisfactory. The yellow catfish industry faces numerous challenges. Firstly, there is an intensified trend of yellow catfish miniaturization, leading to a severe decline in the species due to overfishing of wild stocks [9]. Secondly, inbreeding poses difficulties in ensuring production benefits during the breeding process. The economic traits of the seedlings deteriorate due to varying seedling quality [16,17,18].
A robust line serves is the foundation for aquaculture. The development of breeding varieties with high efficiency, superior quality, and strong disease resistance represents an effective approach to increasing production and efficiency in aquaculture. Across the globe, the aquaculture industry has witnessed significant improvements and advancements owing to breakthrough achievements in improved varieties. Developed countries have recognized improved varieties breeding as a crucial research direction in aquaculture. Examples include rainbow trout bred by the US Dolnelson system [19], Atlantic salmon bred by Chile [20], and whiteleg shrimp (Litopenaeus vannamei), which dominates the world shrimping industry [21]. These new strains have made substantial contributions to global aquaculture production and facilitated the healthy development of the industry. Currently, the emergence of new varieties is urgently needed in Bagridae farming. Selective breeding has become a significant avenue for enhancing fish yield and quality. This discussion focuses on the research progress of Bagridae, considering both conventional breeding technology and advanced breeding technology. Finally, the review summarizes the research progress of hybrid Bagrid catfishes, which represents the mainstream research direction.
3. Conventional Breeding Technology of Bagrid Catfishes
3.1. Selective Breeding
Selective breeding is achieved by carefully selecting male and female animals with reproductive characteristics to reproduce and produce offspring together [22]. Various methods of selective breeding are employed in fish breeding, such as family selection, parent selection, mixed selection, and comprehensive selection [23]. For example, selective breeding has been extensively studied in salmonids [24,25,26,27], cyprinids [28], and channel catfish [29]. Salmon, in particular, has been one of the primary fish species for selective breeding research.
Currently, selective breeding is widely applied in Bagrid catfishes as one of the fundamental methods for selection and breeding. The focus of research in Bagrid catfishes lies mainly in understanding the breeding conditions and the hereditary traits of Pelteobagrus. For instance, a study conducted by Liu et al. [30,31] constructed 63 different families of yellow catfish from Chaohu Lake, Gehu Lake, Hongze Lake, and Taihu Lake as parents. By comparing the growth performance, the Chaohu group was identified as the female parent and the Gehu group as the male parent. In another study, Qin et al. [32] used yellow catfish from five families in Gehu Lake, Shijiu Lake, and Taihu Lake as parents. Through a challenge test involving the injection of Aeromonas hydrophila, the immune performance of different family groups showed significant differences in disease resistance and feed utilization. These studies demonstrate the advantages of selective breeding in enhancing disease resistance, growth rate, and other desirable traits in catfish species.
Selective breeding technology in Pelteobagrus has been predominantly used as the basis for comprehensive breeding technology, with relatively less application in other fishes of the Bagridae family. Therefore, in the subsequent selection process, selective breeding of other catfish species is being explored and integrated into compound breeding technologies to develop new family varieties with improved growth performance.
3.2. Genetic Engineering Applied to Breeding
Genetically modified fish are organisms whose genetic material has been altered using genetic engineering techniques. The primary objective in most cases is to introduce a new trait into the fish species that does not naturally occur, a process known as transgenesis. Genetically modified fish find applications in scientific research, aquaculture, and even as pets [33]. Some genetically modified fish have been engineered to overproduce growth hormone, thereby enhancing their breeding potential. Examples of fish species that have been genetically modified include salmonids [34], carps [35], and tilapias [36].
Lu et al. [37] were the first to obtain transgenic yellow catfish through microinjection. They transferred the genomic DNA of Clarias fuscus into the fertilized eggs of Leiocassis longirostris using DNA microinjection. This approach significantly improved the weight, disease resistance, and hypoxic endurance of Leiocassis longirostris [38]. Ge et al. [39] successfully cloned the growth hormone gene of yellow catfish and established a technical platform for the genome modification of yellow catfish. The mammalian myostatin gene (mstn) serves as a negative regulator in the growth and development of skeletal muscle. In the juvenile yellow catfish, an orthologue of mstna was knocked out using zinc finger nucleases. The resulting genome-edited mstn yellow catfish exhibited normal growth and breeding, with both male and female mutants displaying a distinct muscle phenotype characterized by significant muscular protuberance in the fins after 30 days of post-fertilization. After 80 and 210 days of post-fertilization, the mutants had a higher bodyweight (1.27- to 1.37- fold) compared to wild types [40].
Currently, transgenic technology is still in the early stages of breeding research for Bagrid catfishes. Critics have raised objections to genetically modified fish on various grounds, including ecological concerns, animal welfare concerns, and questions regarding the safety of consuming them as food, as well as the necessity of genetically modified fish to address the world’s food needs [41]. Scientists must carefully weigh the advantages and disadvantages and find solutions to help meet the world’s future needs.
3.3. Cell Engineering Applied to Breeding
Cell engineering applied to breeding involves synthesizing hybrid cells through techniques such as cell culture, nucleic acid induction, nuclear transfer, and cell fusion [42]. This technology can overcome biological reproductive isolation between species, with the main methods being polyploid induction, artificial gynogenesis, and androgenesis [42]. In the case of Bagrid catfishes, the current techniques for cell engineering breeding include artificial gynogenesis and polyploid breeding [43,44,45].
Gynogenesis is an asexual reproduction system that requires the presence of sperm without its DNA actually contributing to the offspring’s genetic material. The paternal DNA dissolves and is destroyed before it can fuse with the egg [46]. Gynogenesis has been observed in various fish species such as Carassius gibelio [47], Oncorhynchus mykiss [48], Paralichthys olivaceus [49], Larimichthys crocea [50], Cyprinus carpio [51], Hypophthalmichthys molitrix [52], Ctenopharyngodon idella [53], Oreochromis mossambicus [54], Megalobrama amblycephala [55], Gobiocypris rarus [56], and Pagrus major [57]. Gynogenetic strains have been successfully developed and utilized in production for Pagrus major and Paralichthys olivaceus [58]. In the case of Bagrid catfishes, pseudo-female fish XY can produce YY supermale yellow catfish through artificial gynogenesis after hormone-induced sex reversal [30,31]. Artificial gonogeny technology is an important component of the technical system for all-male yellow catfish.
Polyploidization of fish aims to modify the genetic makeup of organisms by increasing their genome size [42]. Triploid fish also exhibit strong disease resistance and have delicious meat. Compared to diploid rainbow trout, triploid rainbow trout showed a significantly higher lipid content [59]. Oncorhynchus mykiss triploids and all-female fry culture are prevalent, particularly in Canada [60]. Triploid common carp are approximately one-third thicker than normal common carp. Triploid common carp also possess advantages such as tender and delicious meat, fewer spines, a thick carcass, and a larger body [61]. To breed triploid yellow catfish, triploidy can be achieved through physical or chemical induction of F1 progeny (Figure 5). Various methods such as cold shock, heat shock, hydrostatic pressure rest, and drug treatment have been employed to induce triploidy or tetraploidy in Bagrid catfish [43,44,45,62] (Table 1). It is important to note that different induction methods and treatment temperatures yield varying results. For instance, the triploidization rate and relative emergence rate of yellow catfish embryos were 58% and 53%, respectively, during one induction period [62]. While another induction period yielded a tripling rate of 93.3% and a relative emergence rate of 78.6% [45]. These findings suggest that treatment time is a crucial factor influencing the triploidy of yellow catfish fertilized eggs.
3.4. Molecular Marker-Assisted Breeding
Marker-assisted selection is an indirect selective method used to improve breeding individuals. Currently, molecular marker-assisted breeding is increasingly employed in fish breeding, and common technologies include sequence-related amplified polymorphism (SRAP), randomly amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), simple sequence repeat (SSR), and single nucleotide polymorphism (SNP) [64]. In recent years, the third generation of molecular markers, such as SNP markers, has gained popularity due to advancements in high-throughput sequencing technology. Numerous economic fish species have developed high-density SNP genetic linkage maps and conducted quantitative trait locus (QTL) mapping for various economic traits, including growth [65,66,67,68], sexual maturation [69,70], disease resistance [71,72], and salinity tolerance [69,72].
Currently, most molecular marker development in the Bagridae family focuses on Pseudobagrus ussuriensis and Pelteobagrus fulvidraco (Table 2). Due to the dimorphic characteristics of the Bagridae family, the target traits primarily focus on growth [73,74,75,76] and gender [5,77,78,79,80,81,82,83]. Additionally, markers for stress resistance have also been developed [40]. These markers provide the foundation for molecular-assisted breeding, such as the use of gender markers in the development of new varieties [78]. In our research on P. vachelli, a high-density SNP-based genetic map was developed using the ddRAD-seq technique. Economically important QTL traits were identified, including four weight traits, one body proportion trait, two sex traits, and one hypoxia trait [40]. These results offer a valuable method for the more efficient production of all-male, fast-growing, and hypoxia-tolerant P. vachelli for the aquaculture industry [40].
Overall, research on molecular marker-assisted breeding in the Bagridae family primarily focuses on the gender and growth-related molecular markers of Pseudobagrus ussuriensis and Pelteobagrus fulvidraco, while there are limited reports on other economically important fish species in the Bagridae family, such as Leiocassis longirostris. Furthermore, the development of molecular markers is less related to Bagridae diseases (wheel worm disease, bubble disease, buggy head disease) and stress resistance (hypoxia, temperature) of P. ussuriensis and P. fulvidraco. In the future, it is necessary to develop molecular markers related to other economically important fish species in the Bagridae family and important economic traits. This will effectively clarify the genetic background of Bagridae, help in understand the genetic diversity and structure of the fish, and provide fundamental information for breeding new varieties of Bagridae.
3.5. Hybrid Breeding of Bagrid Catfishes
3.5.1. Interspecific Breeding
Interspecific hybrids are bred by mating individuals from two species, usually within the same genus. In Bagrid catfishes, research on intraspecific hybrids was only found in the genus Pelteobagrus. P. fulvidraco and P. vachelli are the most widely farmed species in the genus Pelteobagrus and have high commercial value [66]. P. vachelli, which is distributed in major Chinese rivers such as the Pearl, Yangtze, Liaohe, Xiangjiang, Huaihe, and Minjiang, is the largest species, with adult individuals reaching a maximum bodyweight of 1850 g and a maximum length of 42.3 cm [84]. On the other hand, P. fulvidraco has a much wider habitat range and is more tolerant to low oxygen levels compared to P. vachelli. However, it reaches a maximum length of 25 cm and a maximum weight of 300 g and exhibits a slower growth rate compared to P. vachelli [85].
Table 3 presents the current status of interspecific breeding in Bagrid catfishes. In reciprocal crosses of P. fulvidraco × P. vachelli, the hybrid combination of P. fulvidraco ♀ × P. vachelli ♂ demonstrates survival rates, fertility rates, and emergence rates exceeding 86.5%, 87.5%, and 71.6%, respectively [86,87,88] On the other hand, the hybrid combination of P. vachelli ♀ × P. fulvidraco ♂ exhibits rates of 79%, 53%, and 6.9% [89]. The cross group displays high heterosis, fast growth rates, and significant absolute weight gain, while the backcross group is unsuitable for production due to low fertilization rates and slow growth [40,90,91,92,93,94,95,96,97,98]. These experimental results lay the foundation for the subsequent acquisition of the hybrid “yellow catfish Huangyou No. 1” from P. fulvidraco ♀ × P. vachelli ♂.
3.5.2. Intergeneric Breeding
Intergeneric hybridization refers to the crossbreeding of fish from different genera within the same family. The family belongs to the branches Chordata, Gridata, Osteichthyes, Actinopterygii, and Siluriformes, which include numerous economically valuable species. For instance, Leiocassis longirostris, which can reach up to 13.0 kg in size, exhibits faster growth compared to P. vachelli (approximately 250 g, with a maximum size of 1.85 kg). Furthermore, L. longirostris possesses higher nutritional value and outstanding meat quality [99,100]. L. longirostris is primarily found in rivers in southern China. Both L. longirostris and P. fulvidraco offer the advantages of delicious meat, fewer muscle spines, and high nutritional value. However, L. longirostris has a larger body size than P. fulvidraco, but its adaptability is inferior, resulting in lower natural yields in water areas [7,88,89]. Pseudobagrus ussuriensis, the largest species among the Bagrid catfishes, is widely distributed in East Asia and is popular among consumers due to its chewy flesh and appealing taste [71]. P. ussuriensis exhibits significant sexual dimorphism in terms of growth rate and body size, with males growing faster than females [101].
In the reciprocal cross of P. vachelli × P. ussuriensis, the length, body weight, absolute weight gain rate, and specific growth rate of the hybrid progeny are higher than those of the parental self-crossing. The gene heterozygosity of hybrids was enhanced, demonstrating a certain degree of heterosis in P. vachelli ♀ × P. ussuriensis ♂ [88,102,103]. Heterosis also occurred in the backcross group, where the absolute weight gain rate and specific growth rate were higher than those of the parental self-crossing. Additionally, the body weight of the hybrid group displayed an advantage over the superparent in P. ussuriensi ♀ × P. vachelli ♂ [88,102]. In reciprocal cross tests of P. fulvidraco × P. ussuriensis, the survival rate, fertility rate, and emergence rate exceeded 88%, 92.67%, and 79.64%, respectively, in the hybrids of P. fulvidraco ♀ × P. ussuriensis ♂, while they were 93%, 89.33%, and 67.33%, respectively, in the hybrids of P. vachelli ♀ × P. fulvidraco ♂ [104,105]. The reciprocal cross results of P. fulvidraco × P. ussuriensis showed little difference, indicating potential worth exploring in the future due to heterosis. In other groups, the appearance and activity distribution were similar to the female parents, but the growth rate was faster than that of the female parents in P. vachelli ♀ × L. longirostris ♂ [106] (Table 4).
In our group, we attempted an intergeneric hybridization. The F1 generation of P. fulvidraco ♀ × L. longirostris ♂ exhibited improved hypoxia tolerance. The orthogonal F1 generation displayed greater genetic diversity and heterozygosity compared to the parents, as determined by SSR molecular marker technology [88]. Furthermore, the orthogonal F1 generation formed a cluster with the male parent L. longirostris on the tree diagram, indicating that the orthogonal F1 generation inherited more genetic material from the male parent [88]. We also investigated the growth conditions of the offspring resulting from the reciprocal cross of P. fulvidraco × L. longirostris. At 30 days, the offspring of P. fulvidraco ♂ × L. longirostris ♀ exhibited different traits. A stubby trait was observed on the right side, contrasting with the normal trait on the left in Figure 6A. Figure 6B depicted a malformed trait with albifaction and hypoplasia of the eyes and mouth. Diversified traits were identified in the offspring of P. fulvidraco ♀ × L. longirostris ♂ at 60 days. Body color became darker in deformed offspring, such as cyclopia (Figure 6D), lack of lateral fin (Figure 6E), and wry mouth (Figure 6F), compared to normal offspring (Figure 6C). After measuring the length and body weight of P. fulvidraco ♀ × L. longirostris ♂ offspring, it was observed that they were higher than those of the offspring from P. fulvidraco ♂ × L. longirostris ♀ after 90 days of growth (unpublished data). Furthermore, the growth rate of the P. fulvidraco ♂ × L. longirostris ♀ offspring was higher than that of the “Huangyou-1” P. fulvidraco ♀ × P. vachelli ♂ offspring after 90 days of measurement. These results indicate that the P. fulvidraco × L. longirostris group has breeding potential in the near future. However, the specific combination with higher breeding value needs to be confirmed by further studies.
According to Table 4, some hybrid generations showed heterosis, but no obvious productive significance was obtained due to a lack of systematic research. Therefore, combining other breeding techniques and leveraging the advantages of hybrids to breed stable hybrid species for market demand is a key breakthrough direction for fish breeding in the Bagridae family in the future.
3.6. Summary and Comparison
Overall, there are relatively few good varieties of the main fish in aquaculture, mainly due to the insufficient connection between basic research and fish genetic breeding practice [58]. Various breeding techniques have their own advantages and disadvantages. By integrating and transferring desirable traits from different varieties or strains using different technical means or combining methods, good varieties can be cultivated. The development of modern biotechnology is crucial for increasing selection accuracy, improving breeding efficiency, and expanding yield. In the future, fish breeding should be improved by combining selective breeding with modern biotechnology, especially molecular genetic markers [23].
4. Research Progress of Hybrid Bagrid Catfishes
4.1. Breeding Conditions
Our team demonstrated that “yellow catfish Huangyou No. 1” can survive in a temperature range of up to 35 °C, with optimal growth observed at a water temperature of 29.8 °C and a stocking density of 1.9 kg/m3. Additionally, water temperature did not affect the sex ratio of P. fulvidraco, P. vachelli, and their hybrid (P. fulvidraco ♀, P. vachelli ♂), with optimum growth temperatures for the three species being 27.53 °C, 28.36 °C, and 28.40 °C, respectively. We initially deduced that the hybrids from P. fulvidraco ♀ × P. vachelli ♂ exhibited heterosis, with a mid-parent heterosis of 22.42% observed during 45 days of breeding juvenile cultivation. Qiang et al. [111] demonstrated that dissolved oxygen concentration (8.3 mg/L) and water temperature (26.0 °C) have optimal synergistic effects on fertilization, hatching, and deformity rates of hybrid yellow catfish (P. fulvidraco ♀ × P. vachelli ♂). It is recommended to maintain the temperature and dissolved oxygen concentration as close as possible to the mentioned values in aquaculture. Even during the hot season, increasing the dissolved oxygen concentration of the water or reducing the temperature by adding fresh water can help improve the breeding efficiency of hybrid yellow catfish. Additionally, there is literature available on supporting breeding technologies, including pond cleaning methods, biological bait culture, methods of introducing fish fries into ponds, taming and feeding techniques, and main disease prevention and control. These resources provide fundamental reference data for the healthy breeding of “yellow catfish Huangyou No. 1” [99].
4.2. Genetic Background
Our team has demonstrated that the hybrids “yellow catfish Huangyou No. 1” were diploids with 52 chromosomes and had the same karyotype formula as their parents [96]. In our laboratory, we cloned the complete mitochondrial genomes of the hybrids (P. fulvidraco ♀ × P. vachelli ♂ and P. fulvidraco ♀ × L. longirostris ♂) and conducted phylogenetic analyses among them [10,96]. It has been determined that the mitochondrial genetic mechanism of the hybrid is maternally inherited [96]. We have systematically investigated the elimination process of paternal mitochondrial DNA and demonstrated that mitochondrial heteroplasmy can affect the embryonic development of hybrid progeny between female P. fulvidraco and male P. vachelli; due to embryonic development, the retention time of paternal mtDNA is later than that of gastral stage [112].
Wang et al. [113] analyzed the spectra of randomly amplified polymorphic DNA (RAPD) bands amplified from the hybrids (P. fulvidraco ♀, P. vachelli ♂) and their parents. The results indicated that the female parent is more closely related to the hybrid offspring. Zhao et al. [114] demonstrated through RAPD analysis that the genetic map of hybrid yellow catfish exhibited three patterns: superposition, superposition-variation, and superposition-weakening. These patterns suggest that the inheritance and heterozygosity of the progeny were significantly enhanced compared to the parents. Additionally, Chen et al. [103] used 18 RAPD random primers to amplify genomic DNA of P. vachelli ♀, P. ussuriensis ♂, and their hybrid offspring (P. vachelli ♀ × P. ussuriensis ♂). They found that the genetic diversity of the hybrid offspring was higher than that of the parents, and the genetic relationship was much closer to the female parent. Most research studies have shown that the genetic relationship of the hybrid progeny is closer to the mother parent [103,113,114]. This genetic information provides the genetic background of heterosis in yellow catfish.
4.3. Molecular Mechanism of Hybrid Vigor
The advent of high-throughput sequencing technology has provided an opportunity to analyze the molecular mechanisms of heterosis in fish, such as Megalobrama amblycephala × Culter alburnus [62], Epinephelus fuscoguttatus × Epinephelus lanceolatus [115], and Takifugu flavidus × Takifugu rubripes [116]. Transcriptome analysis of the hybrid “yellow catfish Huangyou No. 1” and its parents indicated that most of the transcripts determined heterosis in the hybrid yellow catfish. These transcripts were predominantly derived from the parents, but a few deviated from their parental origins [117]. Additionally, the preference for the male parent P. vachelli over the female parent P. fulvidraco in hybrid yellow catfish “Huangyou 1” was due to the significant impact of trans elements on the alleles from the male parent [117].
Our laboratory investigated the key roles of miRNAs in heterosis by integrating transcriptomic, miRNA, and proteomic analyses in hybrid yellow catfish. The results revealed nonadditive expression, homoeologue expression bias, and expression level dominance patterns at the transcriptional, post-transcriptional, and protein levels, providing evidence for the widespread occurrence of dominant models during hybridization. Immune defense, cell proliferation, developmental metabolism, digestion, and absorption were found to be involved in generating the heterosis phenotype. We proposed that high parental gene/protein expression, coupled with low parental miRNAs in the offspring, are inherited from either the mother or father, thereby endowing the offspring with the advantageous traits of either parent and contributing to heterosis.
4.4. Comprehensive Breeding Technology for Bagrid Catfishes
4.4.1. Cell Engineering Breeding and Molecular Marker-Assisted Breeding
Super-male and all-male P. fulvidraco
The manipulation of all-male or all-female populations through genetic methods can greatly benefit fish farming in both theoretical research and production practices [5], Examples include Nile tilapia (Oreochromis niloticus), crucian carp (Carassius carassius), and rainbow trout (Oncorhynchus mykiss). Male and female yellow catfish exhibit significant sexual dimorphism in aquaculture. In full-sibling families, males grow considerably faster than females [5]. Specifically, male yellow catfish displayed a growth rate 130–150% higher than that of females after one year and were nearly twice the size of females after two years [5,30,31]. By employing cell engineering breeding and molecular marker-assisted breeding, the quality, yield, and economic returns of fishery products can be controlled through all-male yellow catfish industries [5,30,31,112]. In 2007, supermales (YY ♀♀) of P. fulvidraco were produced by combining hormone-induced sex reversal with gynogenetic techniques. The combination of sex reversal and progeny testing selection reduced the time required for selective breeding and increased the yield of supermales [31]. Gui et al. [78] developed a method for sex chromosome-specific molecular markers and genetic sex identification in yellow catfish. AFLP molecular marker technology was used to screen and obtain specific AFLP markers for X and Y chromosomes in yellow catfish. By converting them into SCAR markers, a PCR method for sex chromosome genotyping (XX/XY/YY) in yellow catfish was established, with a 100% identification rate [78]. This method was applied to genetically identify the sex of yellow catfish, aiming to produce reliable YY supermales for the production of yellow catfish.
Liu et al. [57] developed a method for sex reversal and gynogenesis to achieve continuous production of yellow catfish. They solved the technical problem by combining the development of female nucleus with hormonal sex reversal to control the sex of yellow catfish. The large-scale breeding technology routes were explored as follows (Figure 7):
Step 1: Sex undifferentiated juvenile fish were obtained. Through hormonal induction techniques of sex reversal, XX and XY physiological female fish were produced via gynogenesis.
Figure 7.
Breeding technology of super-male and all-male yellow catfish P. fulvidraco. Sex undifferentiated juveniles are obtained; through hormonal induction technique of sex reversal, physiological female fish are produced by gynogenesis, and then by sex molecular marker screening, full males are obtained.
Figure 7.
Breeding technology of super-male and all-male yellow catfish P. fulvidraco. Sex undifferentiated juveniles are obtained; through hormonal induction technique of sex reversal, physiological female fish are produced by gynogenesis, and then by sex molecular marker screening, full males are obtained.

Step 2: XY physiological female fish were selected by screening with sex molecular markers and then mated with XY male P. fulvidraco. According to Mendelian law, the quantity ratio of offspring is XX female:XY male:YY male = 1:2:1.
Step 3: Super-male YY fish were obtained by screening with sex molecular markers and mated with normal XX female P. fulvidraco. Finally, all-male XY fish were obtained. The male rate can theoretically reach 100% (in reality, the male rate of progeny of high-quality YY males can reach over 98%) [118].
The novel hybrid species “yellow catfish P. fulvidraco all-male No. 1” has been approved as a new aquatic variety by the National Aquatic Products Original and Improved Seed Approval Committee (Variety Registration No.GS-04-001-2010). In practical production, fish fries of “yellow catfish all-male No. 1” were issued by Wuhan BaiRui Biotech Company (http://fish.job1001.com/company/cm1350436412536/ accessed on 15 June 2020) in 2012. The yield, average size, and breeding benefits of all-male yellow catfish P. fulvidraco were significantly higher than those of normal yellow catfish P. fulvidraco under the same breeding density [119]. “Yellow catfish all-male No. 1” possesses the advantages of normal yellow catfish P. fulvidraco, along with characteristics such as fast growth, neat specifications, a slender and beautiful shape, strong disease resistance, easy feeding, high yield, and high efficiency [120].
4.4.2. Selective Breeding and Hybrid Breeding
Hybrid “yellow catfish Huangyou No. 1” of P. fulvidraco ♀ × P. vachelli ♂
In recent years, the super-male parents have undergone degeneration due to multi-generation self-breeding, resulting in decreased growth and disease resistance in the inbred line. Therefore, farmers have an urgent need to produce new varieties that exhibit faster growth, transport resistance, and disease resistance. Our team, in collaboration with Huazhong Agricultural University, proposed and implemented a technical route that combines selective breeding and hybrid breeding in the creation of the hybrid “yellow catfish Huangyou No. 1” of P. fulvidraco ♀ × P. vachelli ♂ (Figure 8).
Maternal parents were selected from P. fulvidraco populations in four large lakes in the middle reach of the Yangtze River (Liangzi Lake, Hong Lake, Dongting Lake, and Panyang Lake) in 2011. Individuals with unsatisfactory body shape, body color, deformity, and debility were eliminated.
A total of 3800, 3000, 3100, and 3300 individuals were ultimately selected from four different geographic populations. The population of Liangzi Lake was chosen as the foundational population based on a comparative study of growth performance. P. fulvidraco typically achieves sexual maturity within one year. Between 2012 and 2015, three generations of yellow catfish were selected and bred using the following procedures: In 2012, larvae from the F1 generation group of P. fulvidraco were cultivated in Liangzi Lake and subjected to a second selection. In 2013 and 2014, approximately 3 million and 4 million larvae, respectively, were randomly chosen for breeding, resulting in excellent populations of the F2 generation. In 2015, female offspring from the F3 generation group displaying stable growth and were selected for hybridization.
The populations of P. vachelli used for selecting male parents were located in the section of the Yangtze River basin spanning from Yueyang to Wuhan. Undesirable P. vachelli individuals with unsatisfactory body shape, body color, deformities, or weakness were removed. Ultimately, 1800 individuals of P. vachelli were selected as the foundational population. P. vachelli generally reaches sexual maturity in two years. Between 2011 and 2015, two generations of yellow catfish were selected using the following methods: In 2011, larvae from the parental individuals of P. vachelli were randomly selected for cultivation, followed by a second selection. In 2013, approximately 0.8 million larvae were chosen for cultivation, resulting in 2600 premium F1 offspring. In 2015, male offspring from the F2 generation displaying stable growth and were selected for hybridization.
The parent fish (P. vachelli and P. vachelli) were injected with a mixture of spawning-stimulating hormones. Female parents received HCG, LHRH-A2, and DOM, while male parents received half the dosage of HCG, LHRH-A2, and DOM. After an effector phase lasting 14–16 h at a temperature of 24–26 °C, the sperm was preserved in a sperm diluent derived from the testicular homogenate extract of the male parent. Artificial insemination was performed after the eggs were manually extruded. The fertilized eggs were incubated in flowing water at a temperature of 27 °C until hatching, which occurred approximately two days after fertilization. After four days of hatching, the larvae were fed with live cladoceras, rotifers, or Artemia nauplii.
From 2016 to 2018, comparative tests were conducted on the production performance of the hybrid and locally purchased common yellow catfish in Hubei and Jiangsu provinces for two consecutive years. Under the same pond culture conditions and breeding density, the growth rate of the first-instar hybrid, named “yellow catfish Huangyou No. 1”, was 24.17–35.45% faster than that of normal yellow catfish without breeding, and the survival rate was 29.29–33.04% higher.
The novel hybrid species “yellow catfish Huangyou No. 1” has been officially recognized as a new aquatic variety by the National Aquatic Products Original and Improved Seed Approval Committee (Variety Registration No. GS-02-001-2018).
The hybrid species exhibited desirable traits, including high hypoxia tolerance inherited from the female parent P. fulvidraco and strong growth performance from the male parent P. vachelli [95]. Furthermore, the hybrid demonstrated higher nutritive value, disease resistance, and stress tolerance compared to the parental fish.
4.5. Stress Resistance
Currently, there are limited reports on the stress resistance of hybrid yellow catfish. Regarding tolerance to abiotic factors, Wang et al. [113] demonstrated that the oxygen consumption rate of hybrid yellow catfish and P. fulvidraco was significantly lower than that of P. vachelli after 24 h, indicating that the hybrid retained the adaptability of the female parent to lower dissolved oxygen levels. Sun et al. [93] compared the hypoxic tolerance of hybrid yellow catfish and P. fulvidraco under acute hypoxic stress and found that LDH, SOD, CAT, T-AOC activities, and HIF-1α expression levels significantly increased in both hybrid yellow catfish and P. fulvidraco under acute hypoxic stress. Moreover, the hypoxia tolerance of the hybrid yellow catfish was superior to that of P. fulvidraco. Qiang et al. [111] demonstrated that miR-489 targets Copper/Zinc superoxide dismutase to mediate the oxidative response in hybrid yellow catfish (P. fulvidraco ♀, P. vachelli ♂) under Cu2+ exposure. Zhang et al. [40] demonstrated that acyl-CoA thioesterase plays a role in preventing or reducing metabolic disorders in hybrid yellow catfish (P. fulvidraco ♀, P. vachelli ♂) during heat stress in fatty acid metabolism.
Regarding disease resistance, Hu et al. [121] performed a comparative analysis of the half lethal concentration and mortality rate of Edwardsiella ictaluri on P. fulvidraco and hybrid yellow catfish. The mortality rate of the hybrid yellow catfish was significantly lower than that of P. fulvidraco, indicating a significantly improved disease resistance level in the hybrid yellow catfish compared to the female parent P. fulvidraco. Zhang et al. [95] analyzed the survival rates and enzyme activities of copper–zinc superoxide dismutase, manganese superoxide dismutase, and lysozyme after challenging P. fulvidraco, P. vachelli, and their hybrid with Aeromonas hydrophila or Edwardsiella ictaluri. The results indicated that the hybrids possessed a higher survival rate than its parental species after pathogen challenge. The fold change of enzyme activities in the hybrid was higher than that of both parental species after pathogen challenge [95]. The aforementioned research substantiated the obvious heterosis in terms of oxygen tolerance, metal stress tolerance, and disease resistance.
4.6. Metabolic Nutrition
In general, the hybrids exhibited a better nutritional balance than their parents. For example, Shao et al. [117] analyzed the muscle nutritional composition of three breeding populations: P. fulvidraco, P. vachelli, and the hybrid “yellow catfish Huangyou No. 1”. The study found that the crude protein content was 17.55% in P. fulvidraco, 15.89% in P. vachelli, and 18.55% in hybrid yellow catfish, while the crude fat content was 4.15%, 4.56%, and 4.07%, respectively.
This highlights the superiority of hybrids with high protein content and low fat content, as they meet all the indicators set by FAO/WHO in terms of the ratio of total essential amino acids to non-essential amino acids and the proportion of total essential amino acids to total amino acids. Additionally, the hybrids have a rich concentration of fatty acids, indicating that they possess greater nutritional value compared to their parents [117].
Meanwhile, several scholars have conducted research on feed formulation. Zhu et al. [75] indicated that excessive dietary lipid intake can hinder the growth and development of hybrids, leading to imbalances in serum biochemical parameters and a significant reduction in gut microflora diversity. It was found that a normal-fat diet (9% lipids) is more beneficial for the growth of hybrid yellow catfish when compared to a high-fat diet (15% lipids) or a low-fat diet (2% lipids) [75]. Yin et al. [99] and Zhao et al. [100] investigated the effects of dietary isoleucine on the skin mucus barrier and epithelial physical barrier functions of hybrid Bagrid catfish (P. vachelli ♀ × L. longirostris ♂). The results indicated that isoleucine improves the function of the skin mucus barrier by increasing mucus protein and IgM contents, thereby promoting the epithelial physical barrier function in hybrid Bagrid catfish [99,100]. Additionally, the effects of dietary Leucine, threonine, and tryptophan were investigated on various growth performance parameters, including muscle growth and protein synthesis and degradation-related signaling pathways, in hybrid Bagrid catfish (P. vachelli ♀ × L. longirostris ♂) [122,123]. Li et al. [124] explored the growth performance, physiological parameters, and transcript levels of lipid metabolism-related genes in hybrid yellow catfish (P. fulvidraco ♀, P. vachelli ♂) when fed diets containing Siberian ginseng. The results indicated that an appropriate dietary supplementation rate of 2–4 g/kg of Siberian ginseng can influence the regulation of fat metabolism and promote the growth of hybrids [124]. In terms of the mechanism of fatty liver disease, Qiang et al. [125] found that the group with a high dietary lipid level (17.08%) exhibited fatty liver symptoms, such as lower growth, higher hepatic triglyceride and cholesterol contents, and larger lipid droplets in liver tissue, compared to a normal-fat diet group (8.87% dietary lipid level). This was determined through functional miRNA and mRNA regulatory responses. Hybrid yellow catfish demonstrated mediated oxidative degradation of liver glucose and fatty acid peroxidation, which helps protect the liver against stress [125].
5. Summary and Future Perspectives
Improved variety selection involves the application of genetic methods to transform the genetic structure of organisms, aiming to breed high-yield, high-quality, efficient, disease-resistant, and stress-resistant varieties [126]. In recent years, as the study of genetics has progressed, the genetic and breeding techniques of Bagrid catfish have gradually evolved from traditional breeding methods to include cell engineering and marker-assisted selection.
Two new varieties, namely “yellow catfish all-male No. 1” and hybrid “yellow catfish Huangyou No. 1”, have been successfully cultivated in Bagrid catfishes using comprehensive breeding methods. These two new varieties have undergone extensive development for aquaculture.
Table 5 compares the normal yellow catfish with the “yellow catfish all-male No. 1” and hybrid “yellow catfish Huangyou No. 1” in terms of breed type, growth rate, food coefficient, tolerance to low oxygen levels, transportation, specification uniformity, fishing net rate, optimum temperature range, production cost, survival rate, culture cycle, meat quality, thickness of body skin, general disadvantages, and advantages. Both “yellow catfish all-male No. 1” and hybrid “yellow catfish Huangyou No. 1” have the advantage of a short breeding cycle but the disadvantage of high seedling cost. Notably, the hybrid “yellow catfish Huangyou No. 1” exhibits strong resistance to diseases. The male parent of “yellow catfish all-male No. 1” costs RMB 300–500 per unit prize, while the male parent of hybrid “yellow catfish Huangyou No. 1” costs RMB 50–100 per unit price, significantly reducing the seed production cost. Over the past three years, the hybrid “yellow catfish Huangyou No. 1” has experienced rapid development and is gradually replacing the “yellow catfish all-male No. 1” and normal yellow catfish, occupying a dominant position in yellow catfish breeding. Other hybrids, such as P. fulvidraco × L. longirostris, are still in the experimental stage. Although these new varieties of yellow catfish offer certain advantages, farmers still require better breeding techniques, including improved tolerance to low oxygen levels and disease resistance, as well as greater economic value from new yellow catfish strains.
5.1. Whole Genome Sequencing
The genetic mechanism and molecular analysis of important economic traits have become the focal point of current fish research. Since the 21st century, the advent and widespread use of the new generation of SNP typing, and even whole-genome sequencing, have significantly advanced our comprehensive understanding of life sciences, characterized by high throughput and cost-effectiveness [127]. To date, the genomes of model fish have not only been deciphered but also the genomes of several non-model animals have been successively published. For instance, Liu et al. [128] analyzed the molecular mechanism of heterosis in intergenera hybridization (goldfish ♀ × common carp ♂) through genome sequencing. RNA-seq analyses revealed extensive chimeric genes and immediate mutations of orthologs in both diploid and tetraploid offspring. In the case of hybrid Bagrid catfishes, the utilization rate of fish heterosis surpasses our theoretical understanding. To comprehensively explore the molecular genetic mechanism underlying heterosis formation in hybrid yellow catfish at the genomic level, it is imperative to conduct complete genome sequencing of the hybrid “yellow catfish Huangyou No. 1”, as well as P. fulvidraco and P. vachelli. Our team, in collaboration with other units, has sequenced the genomes of “yellow catfish Huangyou No. 1” and P. vachelli based on the published P. fulvidraco genome. Acquiring this series of data will not only provide strong support for further research on the genetic mechanism of heterosis but also accelerate the breeding process of fish genomes as a whole.
5.2. Establishment of Germplasm Resource Bank
During the process of farmers’ management, excellent hybrid strains may inadvertently enter the natural environment due to breeding oversight, potentially impacting the germplasm resources of Bagrid catfish. Additionally, the development of new strains through gene editing technology is progressing. Future exploration of mutated genes with favorable production traits may be necessary. However, these genetic engineering breeding practices have resulted in litigation, international trade disputes, protests, and strict regulations on commercial products in most countries [129].
To address concerns about the contamination of transgenic and hybrid strains [130,131], it is crucial to strengthen breeding management and establish germplasm standards or a germplasm bank for Bagrid catfish. This measure will provide a theoretical foundation for protecting the genetic diversity of Bagrid catfish and promote the sustainable utilization of Bagrid resources, ultimately facilitating the healthy development of Bagrid aquaculture.
5.3. Improving Original New Varieties and Breeding Other Hybrid Bagrid Combinations
Improving existing new varieties can be achieved through selective breeding, hybrid breeding, and molecular marker-assisted breeding. For instance, a genome-wide association study (GWAS) was conducted using the catfish 250 K SNP array to identify quantitative trait loci (QTLs) associated with tolerance to low dissolved oxygen or heat stress in the female channel catfish × male blue catfish interspecific system [132]. These findings have significant implications for understanding genomic evolution in catfish and the integration of hybrid production programs in aquaculture [128,133].
In the case of Bagrid catfish, the all-male hybrid “yellow catfish Huangyou No. 1” can be considered a novel variety for expanded production. With reference to Figure 7, YY P. vachelli and all-male hybrid yellow catfish “Huangyou-1” can be created. However, the lack of gender markers in molecular marker-assisted breeding for these two potential new varieties [40] limits their development. Therefore, completing genome sequencing for these potential new species, as mentioned earlier, is necessary. Building on the widespread adoption and application of the above techniques, it is essential to explore breeding other hybrid parental combinations in Bagrid catfish.
5.4. Problems and Challenges
5.4.1. Genetic Breeding
Genetic breeding is a crucial approach for improving fish yield and quality [134]. However, several issues plague the Bagrid catfish industry. These include inadequate identification and preservation of excellent germplasm resources, insufficient genetic analysis of important traits, and uncertainties surrounding breeding products, such as genetically modified fish. Transgenic technology is still in the early stages of research for Bagrid catfish. Scientists need to carefully consider the advantages and disadvantages of using genetically modified fish to meet global food needs, considering ecological concerns, animal welfare, and the safety of GM food. Therefore, it is necessary to enhance the level of science and technology in genetic breeding and foster independent innovation capabilities.
5.4.2. Broodstock Management
The parents of the Bagridae family are vital carriers of favorable genes and form the basis of breeding; effective management and utilization of parent fish are both essential for promoting the healthy development of the fishery economy. However, there are several challenges in Bagridae management.
Improper selection of parent fish, such as choosing individuals of inappropriate age, vitality, body type, and size, can result in weak disease resistance and growth performance in offspring. When introducing parent fish, it is important to adhere to standard procedures and ensure clear population pedigree, as there are potential concerns related to inbreeding and multi-generation self-breeding. In some cases, certain varieties with geographical characteristics are harvested directly from the wild environment or lack proper analysis of population heritability for the convenience of breeding seedlings. Consequently, these practices lead to a degradation of variety characteristics, loss of advantages, and decline in economic traits. Additionally, there is a lack of genetic information on parent fish germplasm in certain farms, where individual labeling, gene sequencing analysis, and information storage and application are absent.
The aquaculture industry faces various environmental challenges. Increasing the yield of Bagridae fish will require addressing extensive environmental issues. Climate change poses a further threat, necessitating significant reductions in the environmental impact of Bagridae fish for sustainable aquaculture growth [134]. Fish diseases are also a persistent problem, such as pathogen infections, intestinal sepsis, ulcer syndrome, ascites, red head, and split head. These diseases easily affect artificial breeding, resulting in immune dysfunction and reduced production. It is crucial to explore appropriate breeding methods based on the local ecological environment, rational food supply, and disease prevention and treatment. Moreover, the decline of fish resources is a pressing issue, as there is a shallow understanding of resource protection and overfishing of wild fish, leading to biological depletion [134]. In the case of Bagridae fish, overfishing contributes to the miniaturization trend of yellow catfish and a significant reduction in number of species.
Considering the aforementioned problems, it is essential to implement effective measures to alleviate resource scarcity and environmental damage, thereby promoting the development of sustainable and environmentally friendly aquaculture practices.
6. Conclusions
In conclusion, this review highlights progress in the research on the breeding conditions, genetic background, stress resistance, metabolic nutrition, and heterosis of hybrid Bagrid catfish. It provides prospective ideas for the genetic breeding of Bagrid catfish and emphasizes the urgency in yellow catfish production and seed demand in recent years. The review also summarizes various types of hybrid yellow catfish, including “Huangyou-1” of P. fulvidraco ♀ × P. vachelli ♂, “yellow catfish P. fulvidraco all-male No. 1”, and P. fulvidraco × L. longirostris, based on the latest research results from our research group. It further explains the optimal combination for market needs, which is “Huangyou-1” of P. fulvidraco ♀ × P. vachelli ♂”.
Author Contributions
Conceptualization, G.Z. and Y.Z.; Methodology, X.L. (Xinlan Li); Software, Z.L.; Validation, W.L.; formal analysis, H.L.; Investigation, F.L.; Resources, S.Y. and J.J.; Review, G.Z.; Writing—original draft preparation, L.H.; Writing—review and editing, L.H.; Visualization, X.L. (Xia Liang); Supervision, Writing—review and editing, Q.C.; Project administration and funding acquisition, X.L. (Xia Liang) and Q.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was granted by the National Natural Science Foundation of China (32102754, 30770283), Project Foundation of the Academic Program Development of Jiangsu Higher Education Institution (PAPD), Key Research and Development Program of Jiangsu Province (BE2017377), Creation Project of Major New Species of Agriculture in Jiangsu Province (PZCZ201742), and the Natural Science Foundation of Jiangsu Province (No. 18KJB240001), Qingchuang science and technology support program of Shandong provincial college and Natural Science Foundation of Shandong Province of China (ZR2019QC002).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
P. fulvidraco indicated Pelteobagrus fulvidraco; P. vachelli indicated Pelteobagrus vachelli; L. longirostris indicated Leiocassis longirostris; P. ussuriensi indicated Pseudobagrus ussuriensis.
References
- Nelson, J.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Zhang, G.; Deng, S.; Zhang, H.; Li, H.; Li, L. Distribution of different taste buds and expression of α-gustducin in the barbells of yellow catfish (Pelteobagrus fulvidraco). Fish Physiol. Biochem. 2006, 32, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Chen, X.; Zhu, X.; Li, X.; Zhao, Y.; Xu, Z.; Liu, W. Insulin-like growth factor I of yellow catfish (Pelteobagrus fulvidraco): cDNA characterization, tissue distribution, and expressions in response to starvation and refeeding. Fish Physiol. Biochem. 2020, 46, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Gui, J.-F. Genetic basis and biotechnological manipulation of sexual dimorphism and sex determination in fish. Sci. China Life Sci. 2015, 58, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Zheng, M.; Lin, G.; Fang, W.; Huang, J.; Lu, J.; Sun, X. Construction of high-density genetic map and mapping of sex-related loci in the yellow catfish (Pelteobagrus fulvidraco). Mar. Biotechnol. 2020, 22, 31–40. [Google Scholar] [CrossRef]
- Tai, Z.P.; Li, D.D.; Ling, S.C.; Zhang, D.G.; Cui, H.Y.; Tan, X.Y. Identification of full-length cDNA sequences for three development-relevant genes from yellow catfish Pelteobagrus fulvidraco and their transcriptional responses to high fat diet. Comp. Biochem. Physiol. Part B 2018, 225, 67–74. [Google Scholar] [CrossRef]
- Wang, Q.I.; Shen, J.; Yan, Z.; Xiang, X.; Mu, R.; Zhu, P.; Yao, Y.; Zhu, F.; Chen, K.; Chi, S.; et al. Dietary Glycyrrhiza uralensis extracts supplementation elevated growth performance, immune responses and disease resistance against Flavobacterium columnare in yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish. Immunol. 2020, 97, 153–164. [Google Scholar] [CrossRef]
- Li, D.; Xie, C.; He, X.; Tan, R.; Tian, X.; Zhang, Z.; Gao, Y. The Success of Yellow Catfish Aquaculture in China: From Rare Wild Fish to Popular Farmed Fish. In Aquaculture in China: Success Stories and Modern Trends; Wiley: Hoboken, NJ, USA, 2018; pp. 270–282. [Google Scholar]
- Jia, B.; St-Hilaire, S.; Singh, K.; Gardner, I.A. Farm-level returns and costs of yellow catfish (Pelteobagrus fulvidraco) aquaculture in Guangdong and Zhejiang provinces, China. Aquac. Rep. 2016, 4, 48–56. [Google Scholar] [CrossRef]
- Zhang, G.; Tao, P.; Chen, J.; Wang, R.; Zang, X.; Yin, S. The complete mitochondrial genome of the hybrid of Pelteobagrus fulvidraco (female symbol) × Pelteobagrus vachelli (male symbol). Mitochondrial DNA Part A 2015, 27, 4191–4192. [Google Scholar] [CrossRef]
- Liu, J.Y.; Li, A.H.; Zhou, D.R.; Wen, Z.R.; Ye, X.P. Isolation and characterization of Edwardsiella ictaluri strains as pathogens from diseased yellow catfish Pelteobagrus fulvidraco (Richardson) cultured in China. Aquac. Res. 2010, 41, 1835–1844. [Google Scholar] [CrossRef]
- Cheng, K.; Tang, Q.; Guo, X.; Karow, N.A.; Wang, C. High dose of dietary vitamin D3 modulated the yellow catfish (Pelteobagrus fulvidraco) splenic innate immune response after Edwardsiella ictaluri infection. Fish Shellfish. Immunol. 2020, 100, 41–48. [Google Scholar] [CrossRef]
- Cheng, K.; Tang, Q.; Huang, Y.; Liu, X.; Karrow, N.A.; Wang, C. Effect of vitamin D3 on the immunomodulation of head kidney after Edwardsiella ictaluri challenge in yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish. Immunol. 2020, 99, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wei, C.C.; Ye, H.M.; Zhao, H.P.; Song, Y.F.; Wu, K. Effect of dietary choline levels on growth performance, lipid deposition and metabolism in juvenile yellow catfish Pelteobagrus fulvidraco. Comp. Biochem. Physiol. Part B 2016, 202, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.; Li, L.; Guo, S.; Bai, J.; Fedor, C.; Naylor, R.L. Feed and fishmeal use in the production of carp and tilapia in China. Aquaculture 2013, 414–415, 127–134. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, K.Y.; Li, C.W.; Ren, S.Y.; Zhou, Z.Y.; Liu, X.X.; Liu, X.F.; Lai, W.M.; Huang, X.L.; Chen, D.F. Isolation and Characterization of Edwardsiella ictaluri from Southern Catfish, Silurus soldatovi meridionalis, Cultured in China. J. World Aquac. Soc. 2013, 44, 273–281. [Google Scholar] [CrossRef]
- Wang, K.L.; Ji, W.; Zhang, G.R.; Wei, K.J.; Shi, Z.C.; Zhang, X.T.; Zheng, H.; Fan, Q.X. Molecular characterization and expression analysis of three TLR genes in yellow catfish (Pelteobagrus fulvidraco): Responses to stimulation of Aeromonas hydrophila and TLR ligands. Fish Shellfish. Immunol. 2017, 66, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.; Ma, Z.; Li, T.; Xing, W.; Jiang, N.; Li, W.; Li, C.; Luo, L. Effects of totally replacing dietary fish oil by linseed oil or soybean oil on juvenile hybrid sturgeon, Acipenser baeri Brandt♀ × A. schrenckii Brandt♂. Aquac. Nutr. 2018, 24, 184–194. [Google Scholar] [CrossRef]
- Ostberg, C.O.; Chase, D.M.; Hauser, L. Hybridization between yellow-stone cutthroat trout and rainbow trout alters the expression of muscle growth-related genes and their relationships with growth patterns. PLoS ONE 2015, 10, e0141373. [Google Scholar] [CrossRef]
- Lhorente, J.P.; Araneda, M.; Neira, R.; Yáñez, J.M. Advances in genetic improvement for salmon and trout aquaculture: The Chilean situation and prospects. Rev. Aquac. 2019, 11, 340–353. [Google Scholar] [CrossRef]
- Andriantahina, F.; Liu, X.; Huang, H. Genetic map construction and quantitative trait locus (QTL) detection of growth-related traits in Litopenaeus vannamei for selective breeding applications. PLoS ONE 2013, 8, e75206. [Google Scholar] [CrossRef]
- Gjedrem, T.; Baranski, M. Selective Breeding in Aquaculture: An Introduction; Springer: New York, NY, USA, 2009. [Google Scholar]
- Zhou, J.; Ma, A.; Wang, X. Advances in fish breeding techniques. Fish. Inf. Strateg. 2013, 28, 199–207. (In Chinese) [Google Scholar]
- Okamoto, N.; Tayama, T.; Kawanobe, M.; Fujiki, N.; Yasuda, Y.; Sano, T. Resistance of a rainbow trout strain to infectious pancreatic necrosis. Aquaculture 1993, 117, 71–76. [Google Scholar] [CrossRef]
- Thodesen, J.; Grisdale-Helland, B.; Helland, S.J.; Gjerde, B. Feed intake, growth and feed utilization of offspring from wild and selected Atlantic salmon (Salmo salar). Aquaculture 1999, 180, 237–246. [Google Scholar] [CrossRef]
- Kause, A.; Ritola, O.; Paananen, T.; Wahlroos, H.; Mäntysaari, E.A. Genetic trends in growth, sexual maturity and skeletal deformations, and rate of inbreeding in a breeding programme for rainbow trout (Oncorhynchus mykiss). Aquaculture 2005, 247, 177–187. [Google Scholar] [CrossRef]
- Storset, A.; Strand, C.; Wetten, M.; Kjøglum, S.; Ramstad, A. Response to selection for resistance against infectious pancreatic necrosis in Atlantic salmon (Salmo salar L.). Aquaculture 2007, 272, S62–S68. [Google Scholar] [CrossRef]
- Moav, R.; Wohlfarth, G. Two-way selection for growth rate in the common carp (Cyprinus carpio L.). Genetics 1976, 82, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Bondari, K. Response to bidirectional selection for body weight in channel catfish. Aquaculture 1983, 33, 73–81. [Google Scholar] [CrossRef]
- Liu, H.; Guan, B.; Xu, J.; Hou, C.; Tian, H.; Chen, H. Genetic manipulation of sex ratio for the large-scale breeding of YY super-male and XY all-male yellow catfish (Pelteobagrus fulvidraco). Mar. Biotechnol. 2013, 15, 321–328. [Google Scholar] [CrossRef]
- Liu, P.; Chen, X.; Zhong, L. Comparison of growth performance of different families of yellow catfish (Pelteobagrus fulvidraco). Nanjing Norm. Univ. Acta 2013, 36, 90–93. (In Chinese) [Google Scholar]
- Qin, Q.; Wang, M.; Chen, X. Comparative study on immune indexes of different yellow catfish family groups after injection of Aeromonas hydrophila. Freshw. Fish. 2017, 47, 40–46. (In Chinese) [Google Scholar]
- Hackett, P.B.; Alvarez, M.C. The molecular genetics of transgenic fish. Adv. Mar. Biol. 2000, 4, 77–145. [Google Scholar]
- Devlin, R.H.; Vandersteen, W.E.; Uh, M.; Stevens, E.D. Genetically modified growth affects allometry of eye and brain in salmonids. Can. J. Zool. 2012, 90, 193–202. [Google Scholar] [CrossRef]
- Devlin, R.H.; Biagi, C.A.; Yesaki, T.Y.; Smailus, D.E.; Byatt, J.C. Growth of domesticated transgenic fish. Nature 2001, 409, 781–782. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Ronyai, A.; Engidaw, B.Z.; Jauncey, K.; Hwang, G.L.; Smith, A.; Roderick, E.; Penman, D.; Varadi, L.; Maclean, N. Growth and nutritional trials on transgenic Nile tilapia containing an exogenous fish growth hormone gene. J. Fish Biol. 2005, 59, 62–78. [Google Scholar] [CrossRef]
- Lu, J.; Du, W.; Pu, W. Study on breeding technology of transgenic yellow catfish. Sci. Breed Fish 2011, 11, 10. (In Chinese) [Google Scholar]
- Liang, M.; Zhao, B.; You, Y. A preliminary study on the transformation of Clarias fuscus total DNA in catfish long-snout catfish. Southwest China J. Agric. Sci. 1994, 1, 97–101. (In Chinese) [Google Scholar]
- Ge, J.; Song, W.; Dong, Z. Establishment of the first originator of “whole fish” transgenic growth hormone gene in yellow catfish Pelteobagrus fulvidraco. J. Nanjing Univ. Nat. Sci. 2013, 49, 123–131. (In Chinese) [Google Scholar]
- Zhang, G.; Li, J.; Zhang, J.; Liang, X.; Wang, T.; Yin, S. A high-density SNP-based genetic map and several economic traits-related loci in Pelteobagrus vachelli. BMC Genom. 2020, 21, 700. [Google Scholar] [CrossRef]
- Ormandy, E.H.; Dale, J.; Griffin, G. Genetic engineering of animals: Ethical issues, including welfare concerns. Can. Vet. J. 2011, 52, 544. [Google Scholar]
- Zhou, L.; Gui, J. Natural and artificial polyploids in aquaculture. Aquac. Fish. 2017, 2, 103–111. [Google Scholar] [CrossRef]
- Yang, C.; Han, S. Triploid induction of yellow catfish Pelteobagrus fulvidraco. J. Anhui Agric. Sci. 2008, 19, 8118–8119. (In Chinese) [Google Scholar]
- Song, L.; Wang, W.; Zhou, X. Comparative study on triploid induction of yellow catfish (Pelteobagrus fulvidraco) by cold and hot shock methods. J. Fish. 2010, 34, 768–776. (In Chinese) [Google Scholar]
- Li, Y.; Dong, S.; Chen, L. Comparative study on artificial induction of triploid of yellow catfish (Pelteobagrus fulvidraco). J. Tianjin Norm. Univ. 2012, 32, 84–87. (In Chinese) [Google Scholar]
- Avise, J.C. Evolutionary perspectives on clonal reproduction in vertebrate animals. Proc. Natl. Acad. Sci. USA 2015, 112, 8867–8873. [Google Scholar] [CrossRef]
- Gui, J. Artificial propagation and seedling breeding technology of allogynogenetic Carassius auratus gibelio “Zhongke 3”. Rural. Breed. Technol. 2011, 2011, 41–42. (In Chinese) [Google Scholar]
- Sundin, K.; Brown, K.H.; Drew, R.E. Genetic analysis of a development rate QTL in backcrosses of clonal rainbow trout, Oncorhynchus mykiss. Aquaculture 2005, 247, 75–83. [Google Scholar] [CrossRef]
- Gong, C.; Yin, R.; Sun, G. Comparative study on the cultivation of large-scale seedlings of “North Flounder No. 1” and “North Flounder No. 2” and wild flounder. Sci. Breed Fish 2017, 2017, 47–49. (In Chinese) [Google Scholar]
- Xu, J.; You, F.; Wu, X. Artificial induction of diploid gynogenesis in large yellow croaker. Mar. Sci. 2006, 12, 37–42. (In Chinese) [Google Scholar]
- Wu, Q.; Chen, R.; Ye, Y. Study on artificial gynogenesis of carp and its application as a new way to establish inbred line. Genetics 1981, 1, 50–55. (In Chinese) [Google Scholar]
- Mao, H.; Zhou, L.; Gui, J. RAPD analysis of two artificial gynogenetic families of silver carp. Acta Hydrobiol. Sin. 2003, 5, 547–550. (In Chinese) [Google Scholar]
- Zhang, H. Establishment and Main Biological Characteristics of Gynogenetic Grass Carp Population. Ph.D. Thesis, Hunan Normal University, Changsha, China, 2011. (In Chinese). [Google Scholar]
- Yang, N. Studies on Distant Hybridization and Gynogeny of Tilapia (Oreochromis niloticus). Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2020. (In Chinese). [Google Scholar]
- Zhang, X. Study on the Induction of Triploid and Gynogenesis and Its Main Biological Characteristics of Megalobrama amblycephala. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2013. (In Chinese). [Google Scholar]
- Jia, F.; Wang, J.; Wu, Q. Artificial gynogenesis induced by heterologous spermatozoa in Gobio rarei. Acta Hydrobiol. Sin. 2002, 3, 246–252. (In Chinese) [Google Scholar]
- Liu, H.; Hou, J.; Chang, Y. Meiotic gynogenesis of flounder meiosis induced by red bream spermatozoa. J. Fish. 2010, 34, 508–514. (In Chinese) [Google Scholar]
- Xu, K.; Duan, W.; Xiao, J. Application and research progress of biological methods in fish genetics and breeding. Sci. China Life Sci. 2014, 44, 1272–1288. (In Chinese) [Google Scholar]
- Lincoln, R.F.; Scott, A.P. Production of all-female triploid rainbow trout. Aquaculture 1983, 30, 375–380. [Google Scholar] [CrossRef]
- Benfey, T. Use of sterile triploid Atlantic salmon (Salmo salar L.) for aquaculture in New Brunswick, Canada. ICES J. Mar. Sci. 2001, 58, 525–529. [Google Scholar] [CrossRef]
- Cherfas, N.B.; Gomelsky, B.; Ben-Dom, N.; Peretz, Y.; Hulata, G. Assessment of triploid common carp (Cyprinus carpio L.) for culture. Aquaculture 1994, 127, 11–18. [Google Scholar] [CrossRef]
- Wu, C.; Huang, X.; Chen, Q.; Hu, F.; Zhou, L.; Gong, K.; Fu, W.; Gong, D.; Zhao, R.; Zhang, C.; et al. The formation of a new type of hybrid culter derived from a hybrid lineage of Megalobrama amblycephala (♀) × Culter alburnus (♂). Aquaculture 2020, 525, 735328. [Google Scholar] [CrossRef]
- Wu, Q. Study on Artificial Induction of Polyploid of Yellow Catfish (Pelteobagrus fulvidraco). Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2010. (In Chinese). [Google Scholar]
- Tong, J.; Sun, X. Genetic and genomic analyses for economically important traits and their applications in molecular breeding of cultured fish. Sci. China Life Sci. 2015, 58, 178–186. [Google Scholar] [CrossRef]
- Eze, F. Marker-assisted selection in fish: A review. Asian J. Fish. Aquat. Res. 2019, 3, 1–11. [Google Scholar] [CrossRef]
- Wang, C.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y.; Liu, J. Effects of age and dietary protein level on digestive enzyme activity and gene expression of Pelteobagrus fulvidraco larvae. Aquaculture 2006, 254, 554–562. [Google Scholar] [CrossRef]
- Wringe, B.F.; Devlin, R.H.; Ferguson, M.M.; Moghadam, H.K.; Sakhrani, D.; Danzmann, R.G. Growth-related quantitative trait loci in domestic and wild rainbow trout (Oncorhynchus mykiss). BMC Genet. 2010, 11, 63. [Google Scholar] [CrossRef]
- Liu, F.; Sun, F.; Xia, J.H.; Li, J.; Fu, G.H.; Lin, G.; Tu, R.J.; Wan, Z.Y.; Quek, D.; Yue, G.H. A genome scan revealed significant associations of growth traits with a major QTL and GHR2 in tilapia. Sci. Rep. 2014, 4, 7256. [Google Scholar] [CrossRef] [PubMed]
- Küttner, E.; Moghadam, H.K.; Skúlason, S.; Danzmann, R.G.; Ferguson, M.M. Genetic architecture of body weight, condition factor and age of sexual maturation in Icelandic Arctic charr (Salvelinus alpinus). Mol. Genet. Genom. 2011, 286, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.P.; Yáñez, J.M.; Fukui, S.; Swift, B.; Davidson, W.S. Genome-wide association study (GWAS) for growth rate and age at sexual maturation in Atlantic salmon (Salmo salar). PLoS ONE 2015, 10, e0119730. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fan, C.; Liu, Y.; Zhang, Y.; Liu, S.; Sun, D.; Deng, H.; Xu, Y.; Tian, Y.; Liao, X.; et al. A genome scan for quantitative trait loci associated with Vibrio anguillarum infection resistance in Japanese flounder (Paralichthys olivaceus) by bulked segregant analysis. Mar. Biotechnol. 2014, 16, 513–521. [Google Scholar] [CrossRef]
- Zhou, T.; Yuan, Z.; Tan, S.; Jin, Y.; Yang, Y.; Shi, H.; Wang, W.; Niu, D.; Gao, L.; Jiang, W.; et al. A review of molecular responses of catfish to bacterial diseases and abiotic stresses. Front. Physiol. 2018, 9, 1113. [Google Scholar] [CrossRef]
- Zhu, Y.; Liang, H.; Li, Z. Correlation analysis of MSTN gene polymorphism and growth traits in yellow catfish (Pelteobagrus fulvidraco). Genetics 2012, 34, 72–78. [Google Scholar]
- Zhu, C.; Liu, H.; Pan, Z. Construction of a high-density genetic linkage map and QTL mapping for growth traits in Pseudobagrus ussuriensis. Aquaculture 2019, 511, 734213. [Google Scholar] [CrossRef]
- Zhu, C.; Pan, Z.; Wang, H. A Microsatellite Marker and Specific Primers for the Identification of Female and Male Pseudobagrus ussuriensis and Its Application. Patent CN106811540B, 23 June 2020. (In Chinese). [Google Scholar]
- Ge, X.; Yin, H.; Bi, B. Genetic map construction and QTL mapping of growth-related traits in yellow catfish (Pelteobagrus fulvidraco). J. Fish. 2010, 34, 185–193. (In Chinese) [Google Scholar]
- Lu, C.; Sun, X.; Liang, L. Genetic differences between male and female of yellow catfish were analyzed by AFLP. J. Fish. 2007, 20, 24–28. [Google Scholar]
- Gui, J.; Wang, D.; Mao, H. Methods for Sex Chromosome Specific Molecular Markers and Genetic Sex Identification of Yellow Catfish (Pelteobagrus fulvidraco). Patent CN200810236650, 29 April 2009. (In Chinese). [Google Scholar]
- Wang, D.; Mao, H.; Chen, H. Isolation of Y-and X-linked SCAR markers in yellow catfish and application in the production of all-male populations. Anim. Genet. 2009, 40, 978–981. [Google Scholar] [CrossRef]
- Xin, W.; Sun, Z.; Yin, H. SRAP markers of male and female differences in yellow catfish (Pelteobagrus fulvidraco). J. Northeast. For. Univ. 2009, 37, 112–113. (In Chinese) [Google Scholar]
- Xu, H. A Comparative Study on Genetic Diversity among Different Populations of Bagrid (Pseudobagrus ussuriensis). Master’s Thesis, Soochow University, Suzhou, China, 2011. (In Chinese). [Google Scholar]
- Zhu, C.; Pan, Z.; Wang, H. A novel male-associated marker for sex identification in Pseudobagrus ussuriensis. J. Oceanol. Limnol. 2018, 036, 850–852. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, H.; Cheng, L. Identification of sex-specific sequences through 2b-RAD sequencing in Pseudobagrus ussuriensis. Aquaculture 2021, 539, 736639. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, S.; Lv, M. Application of related sequence amplified polymorphism (SRAP) in crop genetics and breeding. Mod. Agric. Sci. Technol. 2010, 11, 79–82. (In Chinese) [Google Scholar]
- Zhang, G.; Yin, S.; Mao, J.; Liang, F.; Zhao, C.; Li, P.; Zhou, G.; Chen, S.; Tang, Z. Integrated analysis of mRNA-seq and miRNA-seq in the liver of Pelteobagrus vachelli in response to hypoxia. Sci. Rep. 2016, 6, 22907. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yan, A.; Zhang, Z.; Zha, J.; Bai, S.; Wu, J. Study on the hybridization of yellow catfish (Pelteobagrus fulvidraco) and yellow catfish (Pelteobagrus vagri). Freshw. Fish. 2002, 03, 3–5. (In Chinese) [Google Scholar]
- Wang, M.B.; Chen, Q.; Chen, Y.B.; Guo, C. Hybridization study of Pelteobagrus fulvidraco and Pelteobagrus vachelli. Mod. Agric. Sci. Technol. 2012, 24, 273–278. (In Chinese) [Google Scholar]
- Wang, F. Morphological difference analysis of F1 generation in Pelteobagrus fulvidraco, Pelteobagrus vagri, Leiocassis longirostris, and their hybrid. Chin. Agric. Bull. 2013, 29, 36–43. (In Chinese) [Google Scholar]
- Wang, F.; Wang, W. Early hybrid breeding of P. fulvidraco, P. vachelli and L. crassilabrus. Aquat. Sci. Technol. Inf. 2004, 1, 10–11. (In Chinese) [Google Scholar]
- Hu, F.; Fan, J.; Qin, Q.; Huo, Y.; Wang, Y.; Wu, C.; Liu, Q.; Li, W.; Chen, X.; Liu, C.; et al. The sterility of allotriploid fish and fertility of female autotriploid fish. Front. Genet. 2019, 10, 377. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Han, L.; Ai, L. Analysis on growth and morphological differences of two hybrid yellow catfish species. Aquaculture 2016, 37, 29–35. (In Chinese) [Google Scholar]
- Pei, X. Studies on Physiological Response and Gene Expression of Hybrid Yellow Catfish “Huangyou 1” to Hypoxia Stress. Master’s Thesis, Nanjing Normal University, Nanjing, China, 2020. (In Chinese). [Google Scholar]
- Sun, J.; Han, G.; Liu, Y. Comparison of growth performance and hypoxia tolerance between hybrid yellow catfish (Pelteobagrus fulvidraco × Pelteobagrus vachelli) and yellow catfish (Pelteobagrus fulvidraco). Acta Hydrobiol. Sin. 2019, 43, 1271–1279. (In Chinese) [Google Scholar]
- Xu, Z.; Zhang, H.; Xu, A. Comparison of breeding of hybrid yellow catfish and all-male yellow catfish. Sci. Fish Farming 2017, 9, 12. (In Chinese) [Google Scholar]
- Zhang, G.; Li, J.; Zhang, J.; Liang, X.; Zhang, X.; Wang, T.; Yin, S. Integrated analysis of transcriptomic, mirna and proteomic changes of a novel hybrid yellow catfish uncovers key roles for mirnas in heterosis. Mol. Cell. Proteom. 2019, 18, 1437–1453. [Google Scholar] [CrossRef]
- Zhang, G.; Liang, X.; Zhang, J.; Li, J.; Yin, S. The complete mitochondrial genome of the hybrid of Leiocassis longirostris (♂) × Pelteobagrus fulvidraco (♀). Mitochondrial DNA Part B 2017, 2, 577–578. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Zhang, G. Microsatellite analysis of genetic diversity of hybrid (P. fulvidraco♀ × P. vachelli♂) and its parents. Fish. Sci. 2018, 37, 612–621. (In Chinese) [Google Scholar]
- Zhou, H.; Duan, G.; Hu, Y.; Wang, H.; Jiang, H.; Ling, J.; Pan, T.; Chen, X. Morphological differences of hybrid yellow catfish with body color differentiation. Fish. Sci. 2021, 40, 1–11. (In Chinese) [Google Scholar]
- Yin, L.; Zhao, Y.; Zhou, X.Q.; Yang, C.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhou, J.; Zhao, J.; et al. Effect of dietary isoleucine on skin mucus barrier and epithelial physical barrier functions of hybrid bagrid catfish Pelteobagrus vachelli × Leiocassis longirostris. Fish Physiol. Biochem. 2020, 46, 1759–1774. [Google Scholar] [CrossRef]
- Zhao, Y.; Yan, M.Y.; Jiang, Q.; Yin, L.; Zhou, X.Q.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, J.; et al. Isoleucine improved growth performance, and intestinal immunological and physical barrier function of hybrid catfish Pelteobagrus vachelli × Leiocassis longirostris. Fish Shellfish. Immunol. 2021, 109, 20–33. [Google Scholar] [CrossRef]
- Pan, Z.J.; Zhu, C.K.; Wang, H.; Chang, G.L.; Ding, H.Y.; Qiang, X.G.; Yu, X.S. Induction of meiotic gynogenesis in bagrid catfish (Pseudobagrus ussuriensis) with homologous sperm and its confirmation for female homogamety. Aquac. Res. 2017, 48, 5659–5665. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, Y.; Chen, X. Morphological difference of F-1 generation in Pelteobagrus vachelli and Pseudobagrus ussuriensis. Fish. Sci. 2011, 23, 264–270. (In Chinese) [Google Scholar]
- Chen, Y.; Chen, X.; Pan, Y.; Wang, M.; Qin, Q.; Deng, Y.; Cai, Y. RAPD analysis of genetic variation of hybrid catfish Pelteobagrus vachelli (♀) × Leiocassis longirostris (♂). J. Shanghai Ocean. Univ. 2010, 19, 12–18. (In Chinese) [Google Scholar]
- Dong, S.; Sun, J.; Bi, X. Changes in body length of P. fulvidraco (♀) × Pseudobagrus ussuriensis (♂) hybrid progeny during embryonic development and 30 days after embryo. Jiangsu Agric. Sci. 2016, 45, 313–315. (In Chinese) [Google Scholar]
- Shi, W.; Chen, Y.; Zhu, C. Study on hybridization effect of P. fulvidraco and P.ussuriensis. Aquac. Res. 2016, 3, 63–70. (In Chinese) [Google Scholar]
- Wei, G. Preliminary study on hybridization in Leiocassis longirostris × Pelteobagrus fulvidraco. Freshw. Fish. 1987, 6, 14–17. (In Chinese) [Google Scholar]
- He, Y.; Zeng, J.; Feng, J.; He, J. Hybrid breeding experiment of P. vachelli (♀) × Leiocassis crassirostris (♂). Aquaculture 2008, 5, 33–34. (In Chinese) [Google Scholar]
- Qiu, C. Seedling breeding and breeding test of P. fulvidraco (♀) × Pseudobagrus ussuriensis (♂). Sci. Technol. Inf. 2009, 4, 313–314. (In Chinese) [Google Scholar]
- Qin, Q.; Liang, D.; Wang, M. The study of embryonic development in Pelteobagrus vachelli and Pseudobagrus ussuriensis. J. Nanjing Norm. Univ. 2012, 35, 81–86. (In Chinese) [Google Scholar]
- Guo, S.; Luo, X.; Liang, H. Mitochondrial DNA sequence of the hybrid of Leiocassis longirostris (♀) and Pelteobagrus fulvidraco (♂). Mitochondrial DNA Part B Resour. 2016, 1, 166–167. [Google Scholar] [CrossRef]
- Qiang, J.; Tao, F.; Bao, W.; He, J.; Liang, M.; Liang, C.; Zhu, H.; Li, X.; Chen, D.; Xu, P. miR-489-3p regulates the oxidative stress response in the liver and gill tissues of hybrid yellow catfish (Pelteobagrus fulvidraco female symbol × P. vachellimale symbol) under Cu (2+) exposure by targeting Cu/Zn-SOD. Front. Physiol. 2019, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Guo, W.; Hu, W.; Mei, J. Delayed elimination of paternal mtDNA in the interspecific hybrid of Pelteobagrus fulvidraco and Pelteobagrus vachelli during early embryogenesis. Gene 2019, 704, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. A Preliminary Study on Hybrid Breeding of P. fulvidraco and P. vachelli. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2003. (In Chinese). [Google Scholar]
- Zhao, W.; Yang, X.; Peng, Z.; Peng, Z.; Liu, Y. Molecular identification of the four species of the genus Pelteobagrus and the genetic analysis of their hybrid. Acta Hydrobiol. Sin. 2006, 30, 101–106. (In Chinese) [Google Scholar]
- Shapawi, R.; Abdullah, F.C.; Senoo, S.; Mustafa, S. Nutrition, growth and resilience of tiger grouper Epinephelus fuscoguttatus × giant Grouper Epinephelus lanceolatus hybrid—A review. Rev. Aquac. 2019, 11, 1285–1296. [Google Scholar] [CrossRef]
- Jiang, H.B.; Bao, J.; Han, Y. Mitochondrial DNA sequence of the hybrid of Takifugu flavidus (♀) × Takifugu rubripes (♂). Mitochondrial DNA Part A 2016, 27, 2117–2118. [Google Scholar] [CrossRef]
- Shao, W. A Preliminary Analysis on the Hybridization Superiority of Yellow Catfish “Huangyou No.1” by Using High-throughput Sequencing Technology. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2018. (In Chinese). [Google Scholar]
- Bao, M.; Feng, J.; Chen, X.; Liu, Z. Artificial propagation technology of all-male yellow catfish (Pelteobagrus fulvidraco). Sci. Fish Breed 2015, 31, 8–9. (In Chinese) [Google Scholar]
- Ye, J.; Dong, T.; Dong, X. Effects of stocking density on yield and benefit of all-male and common yellow catfish (Pelteobagrus fulvidraco) culture. Aquat. Sci. Technol. Inf. 2014, 41, 61–64. (In Chinese) [Google Scholar]
- He, G.; Chen, D. Study on seedling breeding technology of all-male yellow catfish (Pelteobagrus fulvidraco). Hebei Fish. 2016, 2, 37–38. (In Chinese) [Google Scholar]
- Hu, D.; Zhou, Z.; Zhu, Q. Evaluation of resistance ability of hybrid yellow catfish and common yellow catfish to catfish Edwardsiella. Sci. Breed Fish 2016, 2016, 54–55. (In Chinese) [Google Scholar]
- Zhao, Y.; Wu, X.Y.; Xu, S.X.; Xie, J.Y.; Xiang, K.W.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, J.; et al. Dietary tryptophan affects growth performance, digestive and absorptive enzyme activities, intestinal antioxidant capacity, and appetite and GH-IGF axis-related gene expression of hybrid catfish (Pelteobagrus vachelli female symbol × Leiocassis longirostris male symbol). Fish Physiol. Biochem. 2019, 45, 1627–1647. [Google Scholar]
- Zhao, Y.; Jiang, Q.; Zhou, X.Q.; Xu, S.X.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, J.; Jiang, J. Effect of dietary threonine on growth performance and muscle growth, protein synthesis and antioxidant-related signalling pathways of hybrid catfish Pelteobagrus vachelli female symbol × Leiocassis longirostris male symbol. Br. J. Nutr. 2020, 123, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Li, M.X.; Qiang, J.; Bao, J.W.; Tao, Y.F.; Zhu, H.J.; Xu, P. Growth performance, physiological parameters, and transcript levels of lipid metabolism-related genes in hybrid yellow catfish (Tachysurus fulvidraco female symbol × Pseudobagrus vachellii male symbol) fed diets containing Siberian ginseng. PLoS ONE 2021, 16, e0246417. [Google Scholar]
- Qiang, J.; Tao, F.; Bao, W.; He, J.; Li, X.; Chen, J.; Xu, P. Responses of functional miRNA-mRNA regulatory modules to a high-fat diet in the liver of hybrid yellow catfish (P. fulvidraco × P. vachelli). Genomics 2021, 113 Pt 2, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Duan, W.; Xiao, J.; Tao, M.; Zhang, C.; Liu, Y.; Liu, S. Development and application of biological technologies in fish genetic breeding. Sci. China Life Sci. 2015, 58, 187–201. [Google Scholar] [CrossRef]
- Groeneveld, L.F.; Lenstra, J.A.; Eding, H.; Toro, M.A.; Scherf, B.; Pilling, D.; Negrini, R.; Finlay, E.K.; Jianlin, H.; Groeneveld, E.J.A.G.; et al. Genetic diversity in farm animals—A review. Anim. Genet. 2010, 41, 6–31. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Qin, Z.; Geng, X.; Bao, L.; Kaltenboeck, L.; Kucuktas, H.; Dunham, R.; Liu, Z. High-density interspecific genetic linkage mapping provides insights into genomic incompatibility between channel catfish and blue catfish. Anim. Genet. 2016, 47, 81–90. [Google Scholar] [CrossRef]
- Mora, C.; Menozzi, D.; Kleter, G.; Aramyan, L.H.; Valeeva, N.I.; Reddy, G.P. Factors affecting the adoption of genetically modified animals in the food and pharmaceutical chains. Bio-Based Appl. Econ. 2012, 1, 313–329. [Google Scholar]
- Devlin, R.H.; D’Andrade, M.; Uh, M.; Biagi, C.A. Population effects of growth hormone transgenic coho salmon depend on food availability and genotype by environment interactions. Proc. Natl. Acad. Sci. USA 2004, 101, 9303–9308. [Google Scholar] [CrossRef]
- Myhr, A.I.; Dalmo, R.A. Introduction of genetic engineering in aquaculture: Ecological and ethical implications for science and governance. Aquaculture 2005, 250, 542–554. [Google Scholar] [CrossRef]
- Zhong, X.; Wang, X.; Zhou, T.; Jin, Y.; Tan, S.; Jiang, C.; Geng, X.; Li, N.; Shi, H.; Zeng, Q.; et al. Genome-wide association study reveals multiple novel QTL associated with low oxygen tolerance in hybrid catfish. Mar. Biotechnol. 2017, 19, 379–390. [Google Scholar] [CrossRef]
- Jin, Y.; Zhou, T.; Geng, X.; Liu, S.; Chen, A.; Yao, J.; Jiang, C.; Tan, S.; Su, B.; Liu, Z. A genome-wide association study of heat stress-associated SNP s in catfish. Anim. Genet. 2017, 48, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Thompson, S.; Glaser, M. Global aquaculture productivity, environmental sustainability, and climate change adaptability. Environ. Manag. 2019, 63, 159–172. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Species name of Pelteobagrus fulvidraco (A), Pseudobagrus ussuriensis (B), Leiocassis longirostris (C), Pelteobagrus vachelli (D), Tachysurus virgatus (E), and Hyalobagrus ornatus (F) in Bagrid catfishes.
Figure 1.
Species name of Pelteobagrus fulvidraco (A), Pseudobagrus ussuriensis (B), Leiocassis longirostris (C), Pelteobagrus vachelli (D), Tachysurus virgatus (E), and Hyalobagrus ornatus (F) in Bagrid catfishes.

Figure 2.
Research fields of catfish Pelteobagrus according to the analyses of Web of Science (http://apps.webofknowledge.com/ accessed on 15 June 2020) (A); Cited papers related to the catfish Pelteobagrus in past decades (B); Research interests of catfish Pelteobagrus according to analysis of the Carrot database (https://search.carrot2.org/#/search/web accessed on 15 June 2020) (C).
Figure 2.
Research fields of catfish Pelteobagrus according to the analyses of Web of Science (http://apps.webofknowledge.com/ accessed on 15 June 2020) (A); Cited papers related to the catfish Pelteobagrus in past decades (B); Research interests of catfish Pelteobagrus according to analysis of the Carrot database (https://search.carrot2.org/#/search/web accessed on 15 June 2020) (C).

Figure 3.
Yellow catfish aquaculture status in some provinces of China in 2019. (A) indicates different provinces of the People’s Republic of China. Darker color shows higher yellow catfish productions in some provinces. (B) indicates the specific production in the high-ranked provinces. The above data were obtained from the Ministry of Agriculture of the People’s Republic of China (China Fishery Statistical Yearbook, http://english.moa.gov.cn/ accessed on 15 June 2020).
Figure 3.
Yellow catfish aquaculture status in some provinces of China in 2019. (A) indicates different provinces of the People’s Republic of China. Darker color shows higher yellow catfish productions in some provinces. (B) indicates the specific production in the high-ranked provinces. The above data were obtained from the Ministry of Agriculture of the People’s Republic of China (China Fishery Statistical Yearbook, http://english.moa.gov.cn/ accessed on 15 June 2020).

Figure 4.
National yellow catfish seed demand (A) from 2010 to 2019 and production (B) from 2008 to 2019. The above data were obtained from the Ministry of Agriculture of the People’s Republic of China (China Fishery Statistical Yearbook, http://english.moa.gov.cn/ accessed on 15 June 2020).
Figure 4.
National yellow catfish seed demand (A) from 2010 to 2019 and production (B) from 2008 to 2019. The above data were obtained from the Ministry of Agriculture of the People’s Republic of China (China Fishery Statistical Yearbook, http://english.moa.gov.cn/ accessed on 15 June 2020).

Figure 5.
Breeding technology route of triploid yellow catfish. Note: 6-DEMP indicates “6-diethyl methylphosphonate”. F1 was obtained by artificial reproduction, and then triploid yellow catfish was obtained by physical and chemical induction.
Figure 5.
Breeding technology route of triploid yellow catfish. Note: 6-DEMP indicates “6-diethyl methylphosphonate”. F1 was obtained by artificial reproduction, and then triploid yellow catfish was obtained by physical and chemical induction.

Figure 6.
The growth condition of offspring in P. fulvidraco ♂ × Pseudobagrus ussuriensis ♀ after 30 days (A,B). A stubby trait was indicated on the right of (A) compared with the normal trait on the left of (A). (B) indicated a malformed trait with albefaction and hypoplasia of eyes and mouth. The growth condition of offspring in P. fulvidraco ♀ × Pseudobagrus ussuriensis ♂ after 60 days (C–F). Body color turned darker in the deformed offspring such as cyclopia (D), lack of lateral fin (E), and wry mouth (F) compared with the normal traits of offspring (C).
Figure 6.
The growth condition of offspring in P. fulvidraco ♂ × Pseudobagrus ussuriensis ♀ after 30 days (A,B). A stubby trait was indicated on the right of (A) compared with the normal trait on the left of (A). (B) indicated a malformed trait with albefaction and hypoplasia of eyes and mouth. The growth condition of offspring in P. fulvidraco ♀ × Pseudobagrus ussuriensis ♂ after 60 days (C–F). Body color turned darker in the deformed offspring such as cyclopia (D), lack of lateral fin (E), and wry mouth (F) compared with the normal traits of offspring (C).

Figure 8.
Breeding technology route of hybrid yellow catfish “Huangyou 1” P. fulvidraco ♀ × P. vachelli ♂. From 2011 to 2015, P. fulvidraco ♀ and P. vachelli ♂ as parents, the hybrid yellow catfish “Huangyou 1” was obtained through the combination of selective breeding and crossbreeding.
Figure 8.
Breeding technology route of hybrid yellow catfish “Huangyou 1” P. fulvidraco ♀ × P. vachelli ♂. From 2011 to 2015, P. fulvidraco ♀ and P. vachelli ♂ as parents, the hybrid yellow catfish “Huangyou 1” was obtained through the combination of selective breeding and crossbreeding.


{kind=link}
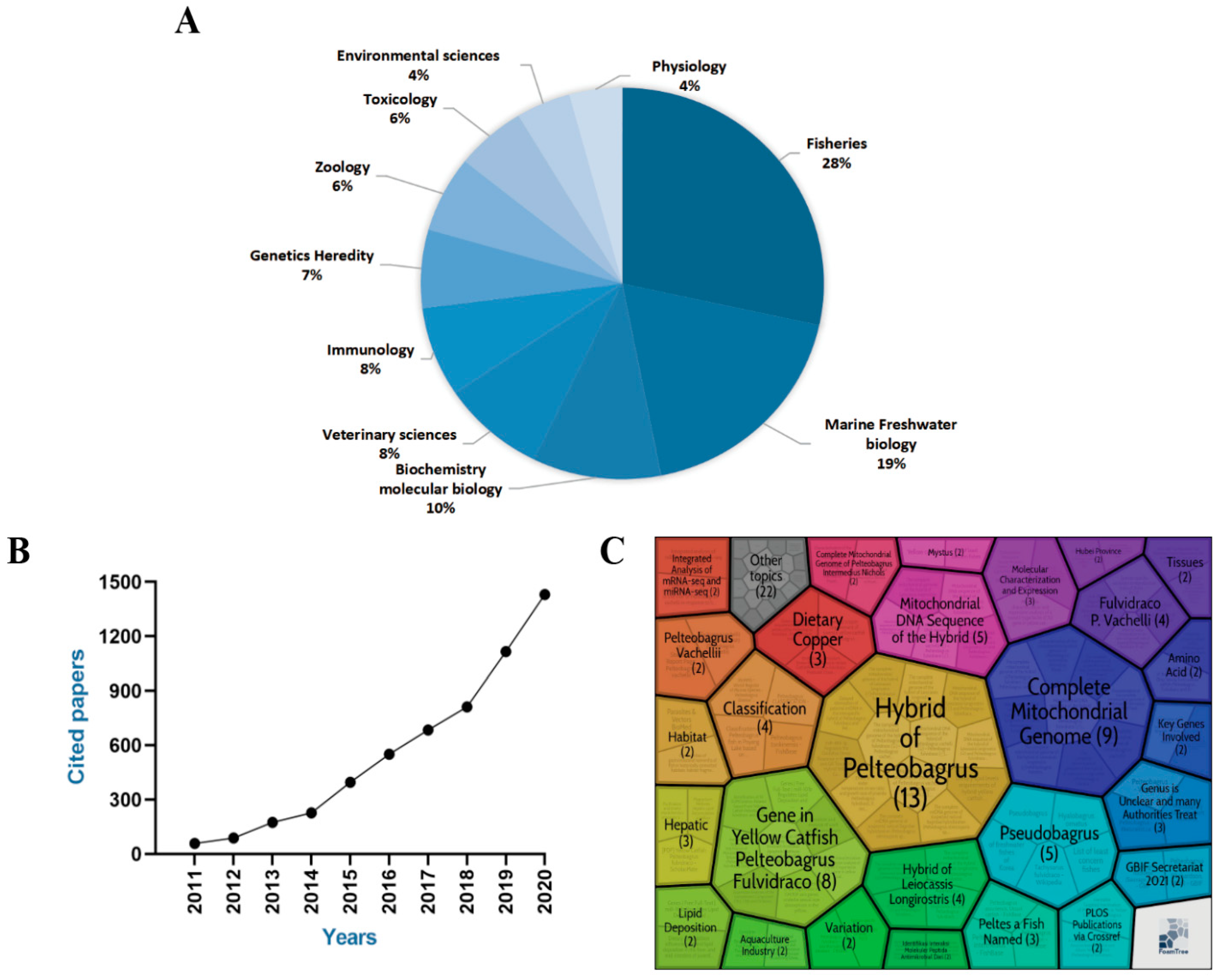{kind=link}
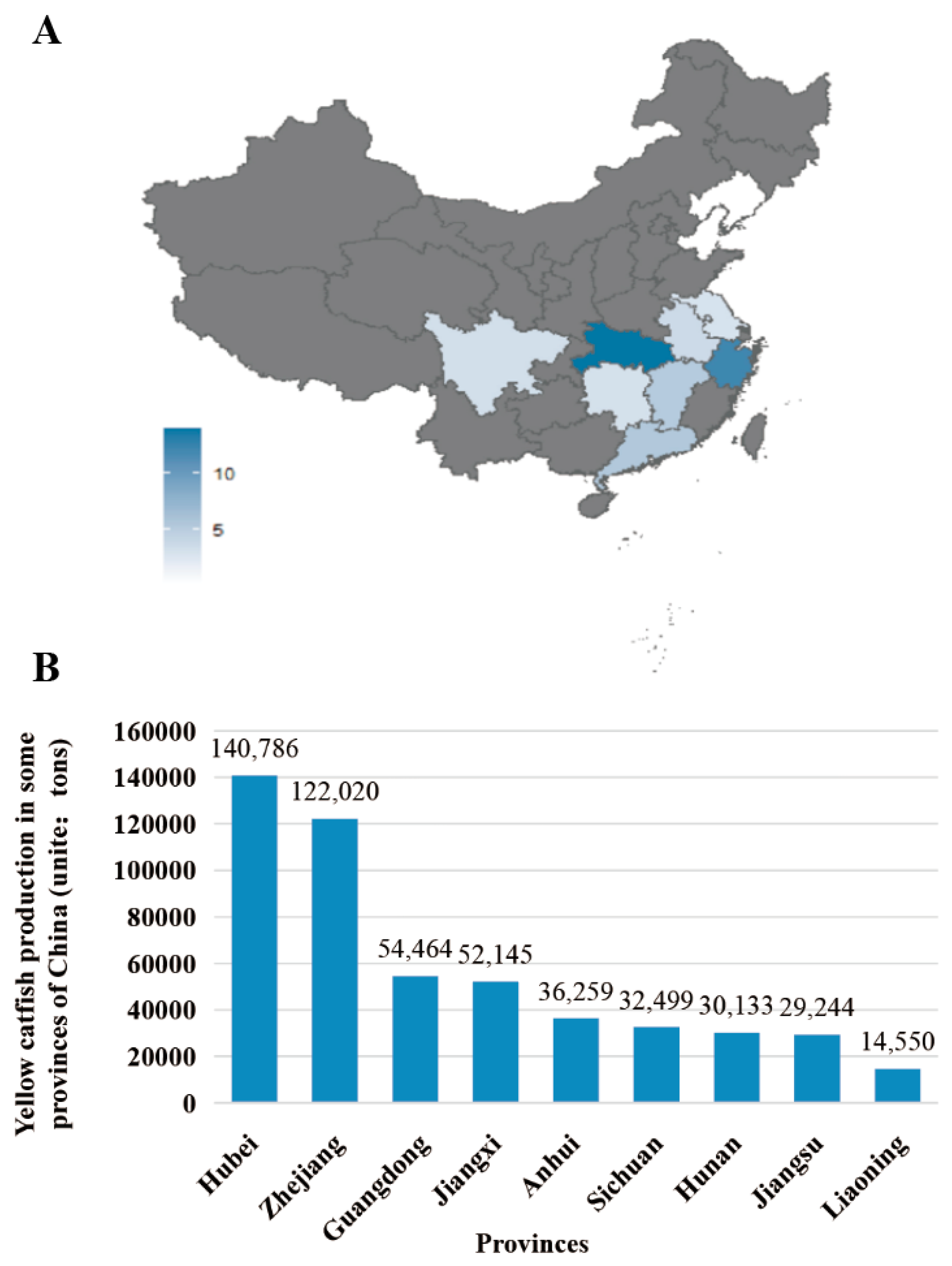{kind=link}
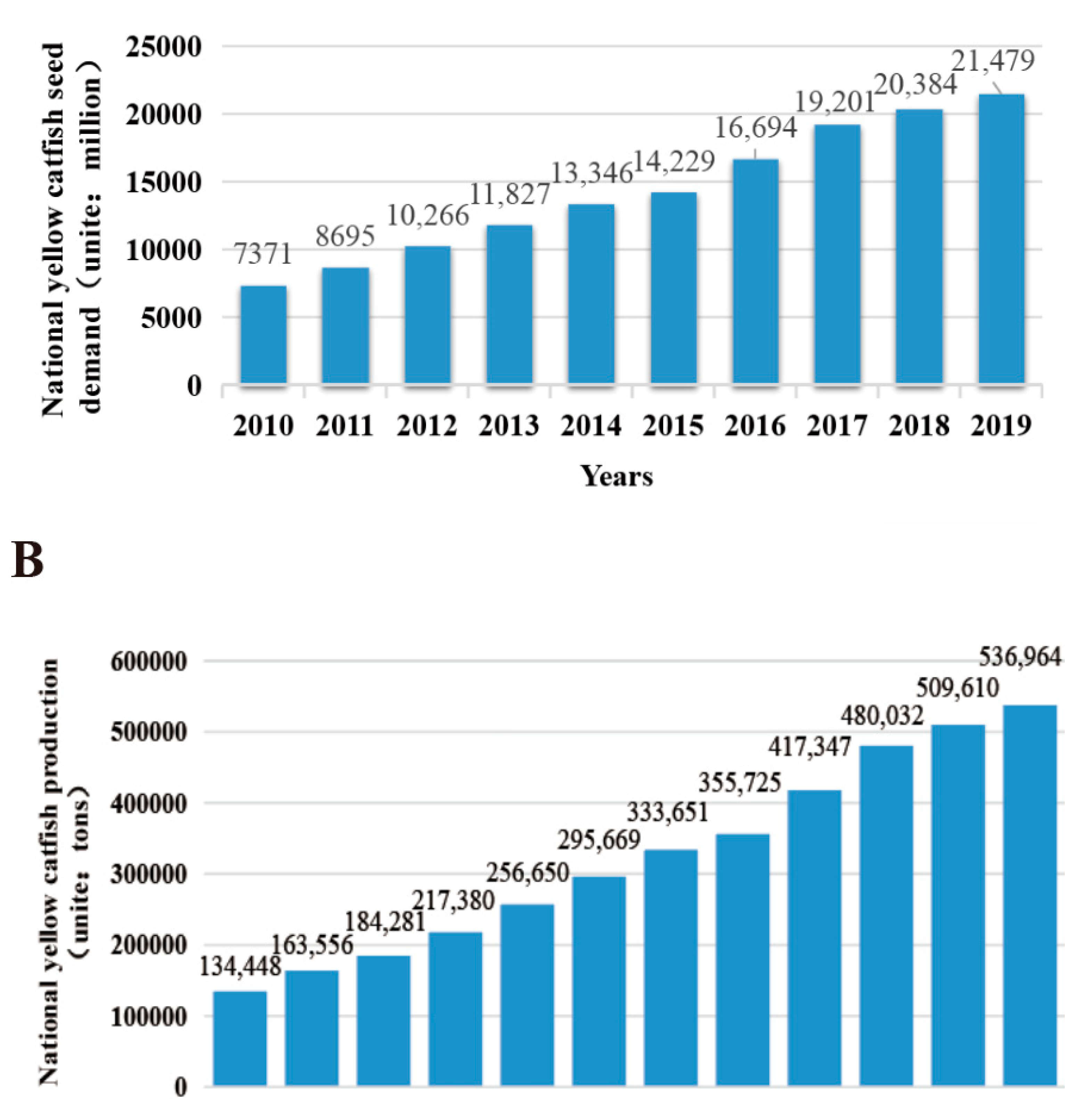{kind=link}
{kind=link}
{kind=link}
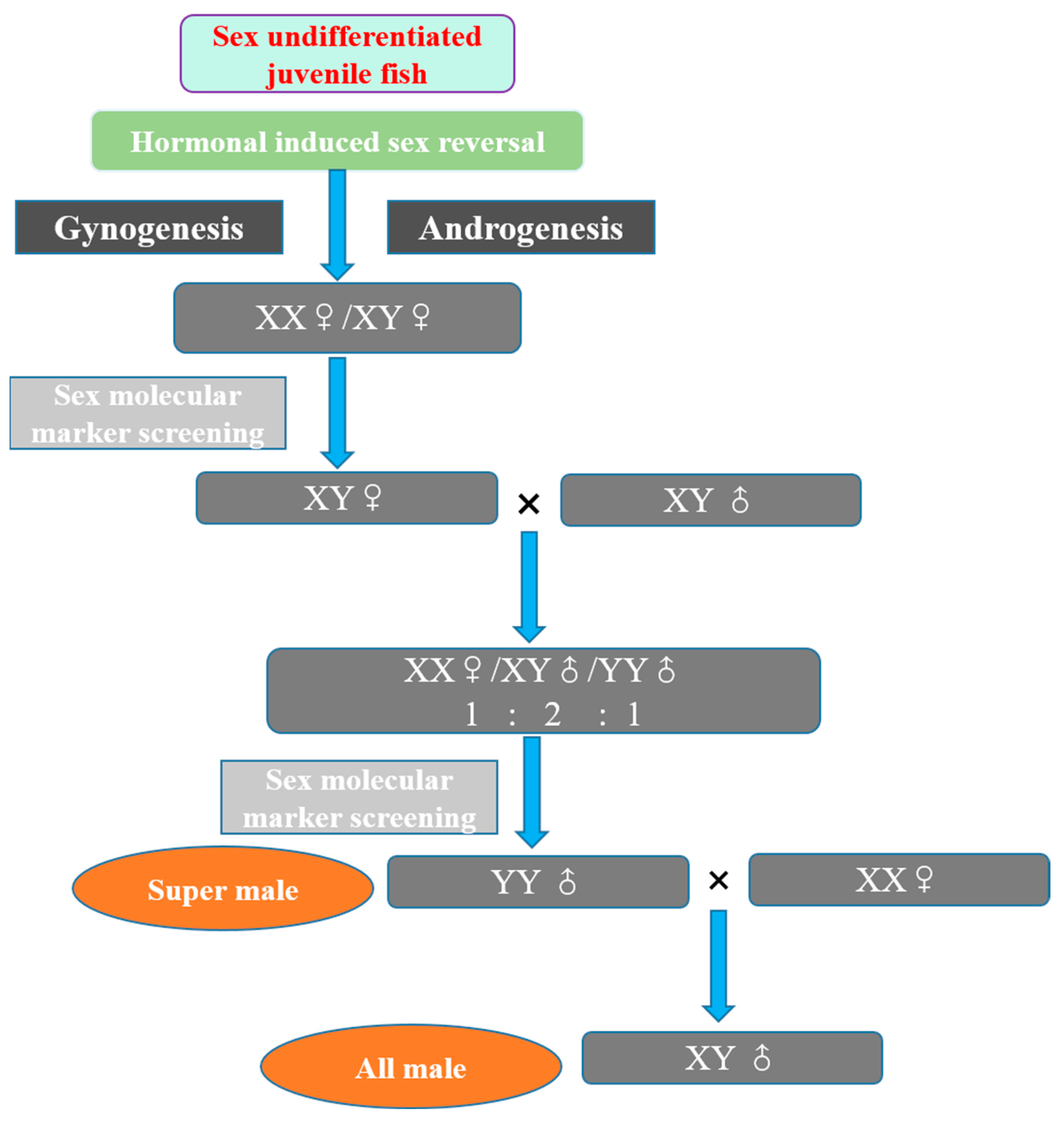{kind=link}
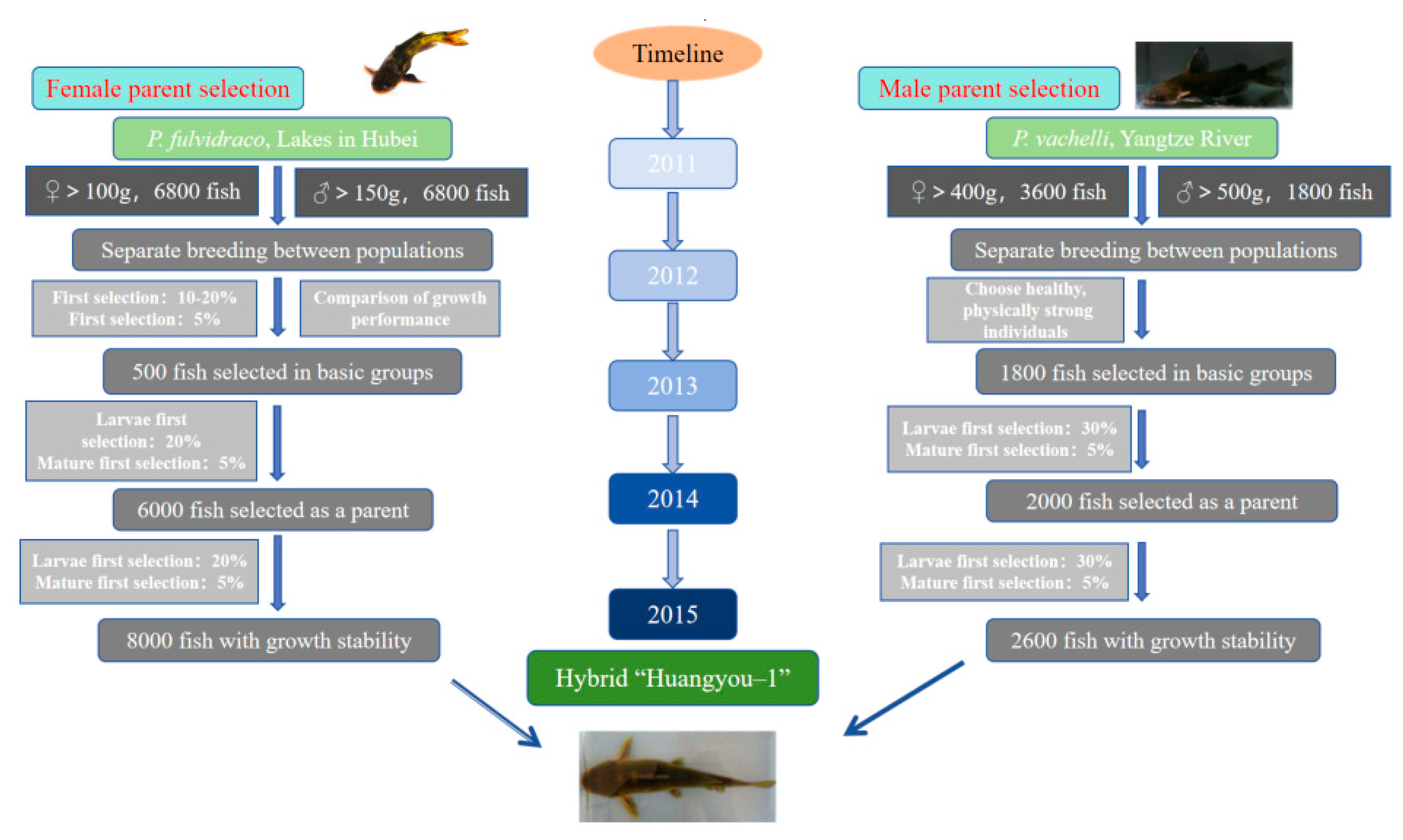{kind=link}
Table 1.
Present situation of polyploid breeding techniques of Bagrid catfishes.
| Authors Year | Species | Polyploid | Inducing Methods | Dose or Temperature | Processing Time | Duration | Results |
|---|---|---|---|---|---|---|---|
| Wu et al., 2010 [63] | P. fulvidraco | Triploid | Cold shock | 4 °C | 3 min after fertilization | 10 min | The triploid and relative emergence rates of embryos were 58% and 53%, respectively |
| Li et al., 2012 [45] | P. fulvidraco | Triploid | Cold shock | 4 °C | 3 min after fertilization | 15 min | Triploid induction rate was 63.3% and hatching rate was 82.3% |
| Song et al., 2010 [44] | P. fulvidraco | Triploid | Cold shock | 5 °C | 2 min after fertilization | 20 min | High deformity rate and mortality for fish fry |
| Song et al., 2010 [44] | P. fulvidraco | Triploid | Hot shock | 40 °C | 2 min after fertilization | 2 min | Triploid induction rate was as high as 60% in embryo and 40~50% in juvenile |
| Wu et al., 2010 [63] | P. fulvidraco | Triploid | Hot shock | 40.5 °C | 2 min after fertilization | 2 min | The triploid rate and relative emergence rate of embryos were 59% and 36%, respectively |
| Wu et al., 2010 [63] | P. fulvidraco | Tetraploid | Hot shock | 40.4 °C | 50 min after fertilization | 1.5–1.8 min | The tetraploid rate of embryos was 10.61–62.16%, the tetraploid rate of embryos in the membrane stage was 11.11–75%, and the relative emergence rate was 9.77–36.05% |
| Li et al., 2012 [45] | P. fulvidraco | Triploid | Hot shock | 40 °C | 8 min after fertilization | 2 min | Triploid induction rate was 93.3% and hatching rate was 78.6% |
| Li et al., 2012 [45] | P. fulvidraco | Triploid | Medication | 6—DMAP, CB | - | - | High triploid deformity rate, low hatching rate, low ploidy rate, not suitable for production |
| Yang et al., 2008 [43] | P. fulvidraco | Triploid | Medication | 6—DMAP | 10 min after fertilization | 15 min | Hatching rate is 0, not suitable for production |
| Wu et al., 2010 [63] | P. fulvidraco | Triploid | Hydrostatic shock | 550 kg/cm2 pressure | 5 min after fertilization | 5 min | Triploid rate and relative emergence rate of embryos were 55% and 54%, respectively |
Note: “-” indicates that the related research results have not been reported. 6-DMAP indicates “6-Dimethylaminopurine”, and CB indicates “cytochalasin B”.
Table 2.
Present situation of molecular marker-breeding of Bagrid catfishes.
| Authors Year | Species | Technologies | Correlated Traits | Research Contents | Results |
|---|---|---|---|---|---|
| Xu, 2011 [81] | P. ussuriensis | SRAP | Gender | The polymorphism of male and female genomic DNA was detected, and their gender related molecular markers were screened | Twelve pairs of primer combinations with clear bands, strong reproducibility, and good polymorphism were screened out |
| Zhu et al., 2018 [82] | P. ussuriensis | PCR; Sequencing | Gender | A total of 95 female and 90 male individuals were examined | A male-associated marker pUGT54 was isolated |
| Zhu et al., 2019 [74] | P. ussuriensis | 2b-RAD | Growth | A high-resolution genetic linkage map was constructed using 2b-RAD technique | A set of QTLs related to growth, body weight, head length, and body width were identified |
| Zhu et al., 2021 [83] | P. ussuriensis | 2b-RAD | Gender | The DNA sequences associated with restrictive endoscopic sites were sequenced to identify sex-specific sequences in 5 female and 5 male fish | Three male-specific markers and seven male-specific sequences were screened |
| Zhu et al., 2020 [75] | P. ussuriensis | PCR | Growth | Selection of sex-specific microsatellite loci in the microsatellite enrichment library | A sex-specific microsatellite locus was selected |
| Zhu et al., 2012 [73] | P. fulvidraco | PCR-SSCP | Growth | SNP detection and MSTN gene were conducted and the association between MSTN gene and growth shape was analyzed | The AA genotype was the favorable genotype and the DD genotype was the unfavorable genotype for the growth |
| Ge et al., 2010 [76] | P. fulvidraco | SSR, SRAP, TRAP | Growth | A genetic linkage map was constructed for QTL mapping of five growth-related traits | All three QTLs could be used for marker-assisted breeding for growth traits |
| Lu et al., 2007 [77] | P. fulvidraco | AFLP | Gender | Genetic differences between males and females were analyzed using 20 AFLP primer combinations | A total of 6 loci showed significant amplification differences between male among 4 AFLP primer combinations |
| Gui et al., 2009 [78] | P. fulvidraco | AFLP | Gender | The PCR method of chromosome genotype identification was established, and it played an important role in production of all-male yellow catfish | Two specific AFLP fragments that could produce X or Y chromosomes were screened and transformed into SCAR markers for sequencing |
| Wang et al., 2009 [79] | P. fulvidraco | AFLP | Gender | 256 pairs of AFLP primers were used to scan the gene pool of XX, XY, and YY | Four X linkage markers and two Y linkage markers were screened out |
| Xin et al., 2009 [80] | P. fulvidraco | SRAP | Gender | A SRAP-PCR amplification reaction system was established, which can be widely used in the genetic study of yellow catfish | One SRAP marker was found to be sex-specific in males and females |
| Gao et al., 2020 [5] | P. fulvidraco | SNP | Gender | Genotyping sequencing was constructed, and 5705 SNP markers were mapped to 26 different linkage groups | 11 significant related QTLs were identified and 6 sex-related genes were identified |
| Zhang et al., 2020 [40] | P. vachelli | SNP | Anti-stress | A high-resolution linkage map was constructed using 5059 DDRAD markers | Candidate genes for 12 growth, 1 sex determination, and 1 hypoxia tolerance related traits were constructed |
Table 3.
Current situation of interspecific breeding in Bagrid catfishes.
| Authors Year | Hybridized Combination | Survival Rate | Fertility Rate | Emergence Rate | Characteristics |
|---|---|---|---|---|---|
| Wang et al., 2002, 2012 [86,87] | P. Fulvidraco ♀ × P. vachelli ♂ | - | 87.5% | 71.6% | High hatching rate and fertilization rate, body color and shape similar to P. fulvidraco, and growth rate is higher than parents |
| Wang et al., 2004 [89] | P. vachelli ♀ × P. fulvidraco ♂ | 79% | 53% | 6.9% | The hybrid progeny had obvious maternal benefit, low fertilization rate, and slow growth rate, which was only 0.1144 g/d and was not suitable for production |
| Wang et al., 2013 [88] | P. fulvidraco ♀ × P. vachelli ♂ | 86.5% | 70.6% | 60.7% | The hybrid combination showed high heterosis, fast growth rate, absolute weight gain rate of 0.1534 g/d, and obvious maternal effect |
| Li et al., 2016 [91] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | Growth speed is fast, and hybrid has the maternal effect |
| Xu et al., 2017 [94] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | The hybrid progeny was superior to all-male yellow catfish in terms of species size and survival rate |
| Zhang et al., 2017, 2018 [96,97] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | The optimal growth temperature and density of hybrid progeny were 28.95 °C and 1.927 kg/m³. Compared with the parents, the progeny had obvious growth advantages, high quality nutritional value, higher genetic diversity, and stronger disease resistance |
| Hu et al., 2019 [90] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | Growth performance was significantly better than P. fulvidraco, and the phenomenon of hermaphroditic growth was significantly reduced. The female was completely sterile, and the sperm quantity in the testis of the male was low and the effective vitality was low |
| Sun et al., 2019 [93] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | The hypoxic tolerance, average body weight, and survival rate were significantly higher than the female parent, and the feed conversion ratio was lower than the female parent |
| Zhang et al., 2019 [95] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | Heterosis in immunity, metabolism, digestion, absorption, proliferation, and development |
| Pei et al., 2020 [92] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | Hypoxic stress induces the upregulation of oxygen-sensing protein-related genes in the progenies of the hybrids and enhances the body’s antioxidant capacity to cope with oxidative stress injury caused by reoxygenation |
| Zhang et al., 2020 [40] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | The hybrid progeny showed heterosis in growth, antimicrobial resistance, and nutrient composition |
| Zhou et al., 2021 [98] | P. fulvidraco ♀ × P. vachelli ♂ | - | - | - | The hybrid progeny showed significant differences in spine length, caudal stalk length, and eye diameter, and the main morphological characteristics of the progeny were more similar to female parent |
Note: “-” indicates that the related research results have not been reported.
Table 4.
Current situation of intergeneric breeding in Bagrid catfishes.
| Authors Year | Hybridized Combination | Survival Rate | Fertility Rate | Emergence Rate | Characteristics |
|---|---|---|---|---|---|
| Wei et al., 1987 [106] | P. Vachelli ♀ × L. longirostris ♂ | - | - | - | The hybrid progeny is between the two parents in shape and grows faster than female parents |
| Wang et al., 2004 [89] | P. Fulvidraco ♀ × L.crassilabrus ♂ | 74% | 64.6% | 11.0% | The hybrids were not suitable for production because of their low emergence rate and morphological characteristics tending to male parents |
| He et al., 2008 [107] | P. Vachelli ♀ × L. longirostris ♂ | - | 70.0% | - | Appearance, activity distribution, and body color similar to P. vachelli |
| Qiu, 2009 [108] | P. Fulvidraco ♀ × P.ussuriensis ♂ | 87% | - | - | The weight gain of hybrid offspring is significantly higher than P. ussuriensis, and there is no significant difference with P. fulvidraco |
| Chen et al., 2010 [103] | P. Vachelli ♀ × P.ussuriensis ♂ | - | - | - | The gene heterozygosity of the first generation of hybridization was enhanced, showing some heterosis |
| Cai et al., 2011 [102] | P. Vachelli ♀ × P. ussuriensis ♂ | - | - | - | The length and body weight of the hybrid progenies were greater than that of the backcrossing and self-crossing progenies |
| Cai et al., 2011 [102] | P.ussuriensis ♀ × P. vachelli ♂ | - | - | - | The body weight of the hybrid group had the superparent advantage |
| Qin et al., 2012 [109] | P.ussuriensis ♀ × P. vachelli ♂ | - | - | - | The embryo development time was between the two parents, and the full length of the hybrid was closer to female parent |
| Wang et al., 2013 [88] | P. Vachelli ♀ × L.crassilabrus ♂ | 76.5% | 81.0% | 72.9% | High fertilization rate and seedling emergence rate, fertilized eggs and strong larvae resistant to high temperature |
| Wang et al., 2013 [88] | P. Vachelli ♀ × P.ussuriensis ♂ | 59.5% | - | - | The absolute weight gain rate and specific growth rate of hybrid progeny are higher than that of self-crossing and lower than that of backcrossing |
| Wang et al., 2013 [88] | P.ussuriensis ♀ × P. vachelli ♂ | 62.5% | - | - | The absolute weight gain rate and specific growth rate of hybrid progeny were higher than that of parental self-crossing |
| Dong et al., 2016 [104] | P. Fulvidraco ♀ × L. longirostris ♂ | - | 92.67% | 79.67% | Earlier embryo development than the female parent. No significant difference in the length of self-mating body between fry and yellow catfish. |
| Guo et al., 2016 [110] | L. Longirostris ♀ × P. fulvidraco ♂ | - | - | - | The genetic distance is closer than that of both parents |
| Dong, 2016 [104] | P. Fulvidraco ♀ × P.ussuriensis ♂ | - | 92.67% | 79.67% | The embryo development was earlier than the female parent and later than the male parent. There was no significant difference in the body length of self-crossing with the female parent |
| Shi, 2016 [105] | P. Fulvidraco ♀ × P.ussuriensis ♂ | 88% | - | - | The fertility advantage of hybrid progeny was not significant |
| Li, 2016 [91] | P. Fulvidraco ♀ × P.ussuriensis ♂ | - | - | - | The hybrid population deviated from the all-male yellow catfish population, and the absolute weight gain rate was higher than the all-male yellow catfish population |
| Shi, 2016 [104] | P.ussuriensis ♀ × P. fulvidraco ♂ | 93% | - | - | The average weight advantage decreased with the increase in time |
| Dong, 2016 [104] | P.ussuriensis ♀ × P. fulvidraco ♂ | - | 89.33% | 67.33% | The embryo development was approximately 14 h shorter than the male parent self-crossing group. The body length was significantly longer than the female parent self-crossing group |
Note: “-” indicates that the related research results have not been reported.
Table 5.
The comparison of cultured yellow catfish, all-male yellow catfish, and hybrid yellow catfish.
Table 5.
The comparison of cultured yellow catfish, all-male yellow catfish, and hybrid yellow catfish.
| Cultured Yellow Catfish | Hybrid Yellow Catfish | All-Male Yellow Catfish | |
|---|---|---|---|
| Breed type | Artificial (high proportion of close relatives) | The female of hybrid fish is sterile | YY crosses with XX |
| Growth rate | The adult male is 30% larger than the female, reaching 100~150 g at the second instar | The average growth rate of yellow catfish was 29.93% faster than that of yellow catfish | 30% faster than the average growth rate of normal yellow catfish |
| Food coefficient | 1.46~1.49 | 1.1~1.3 | 1.1~1.3 |
| Tolerance to low oxygen levels | 0.255 mg/g/h, Choke point = 0.314 mg/mL | 0.206 mg/h, Choke point = 0.27 mg/mL | 0.255 mg/g/h, Choke point = 0.314 mg/mL |
| Transportation | Not tolerant to long distance transportation | Tolerance to long distance transport | Tolerance to long distance transport |
| Specification uniformity | None specification (adult male 30% larger than female) | Specification | Specification (more than 94% of the individual size reached more than 100 g) |
| Fishing net rate | Low | High | Low |
| Optimum temperature range | 22~28 °C and 23~30 °C (breeding water temperature) | 20~32 °C and 28.4 °C (Optimum growth temperature) | 22~28 °C and 23~30 °C (breeding water temperature) |
| Production cost | Low | High | High |
| Survival rate | 33~52% | 89.9~90.3% | 53.3~70.8% |
| Ovulation ratio | 73.6% | - | - |
| Fertilization rate | 72.7% | 79.8% | >80% |
| Hatching rate | 76.8% | 82.5% | 76% |
| Culture cycle | Long | Short | Short |
| Meat quality | High fat content and meat content is 66.47~68.41% | Meat content is 76.52% | High fat content and meat content is 66.47~68.41% |
| Thickness of body skin | Thin | Thick | Thick |
| Disadvantages | Long breeding cycle and low disease resistance | Higher cost of seedlings and a certain rate of malformation | Low survival rate of splashing and high seedling cost |
| Advantages | Lower production cost and lower fry price | Short breeding cycle and strong resistance to disease | Short breeding cycle |
Note: Data of natural yellow catfish is unknown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, L.; Zhang, G.; Zhang, Y.; Li, X.; Luo, Z.; Liu, W.; Luo, F.; Liu, H.; Yin, S.; Jiang, J.; et al. Profiling Genetic Breeding Progress in Bagrid Catfishes. Fishes 2023, 8, 426. https://doi.org/10.3390/fishes8080426
AMA Style
Huang L, Zhang G, Zhang Y, Li X, Luo Z, Liu W, Luo F, Liu H, Yin S, Jiang J, et al. Profiling Genetic Breeding Progress in Bagrid Catfishes. Fishes. 2023; 8(8):426. https://doi.org/10.3390/fishes8080426
Chicago/Turabian StyleHuang, Lishi, Guosong Zhang, Yupeng Zhang, Xinlan Li, Zhong Luo, Wenyu Liu, Fu Luo, Haifeng Liu, Shaowu Yin, Jun Jiang, and et al. 2023. "Profiling Genetic Breeding Progress in Bagrid Catfishes" Fishes 8, no. 8: 426. https://doi.org/10.3390/fishes8080426