
Macro-Habitat Suitability for Threespine Stickleback (Gasterosteus aculeatus L.) Near the Southern Limit of Its Global Distribution: Implications for Species Management and Conservation

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
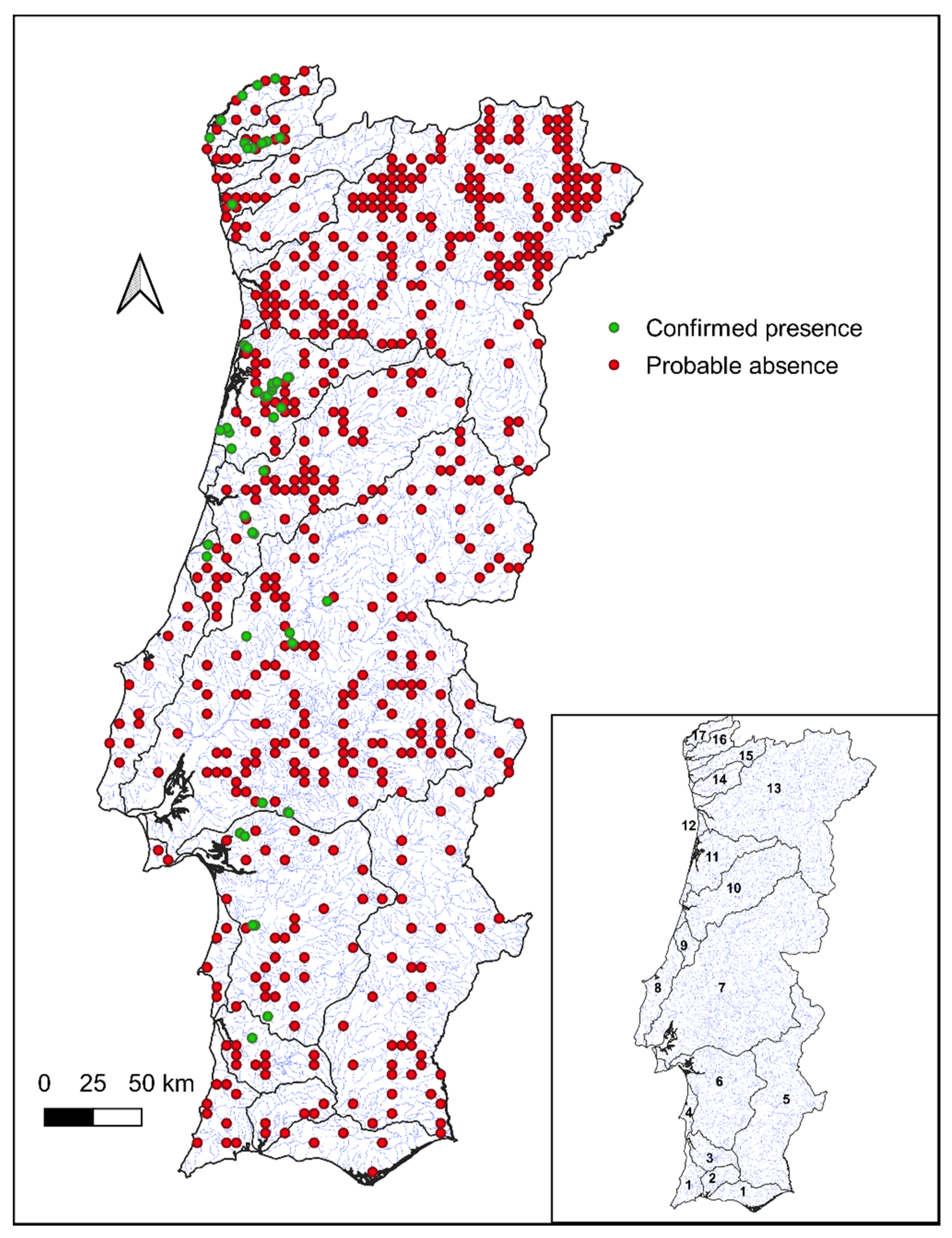
2.2. Species Occurrence Data
2.3. Environmental Predictors
2.4. Statistical Modeling
2.5. Definition of Conservation Priorities
- Level 0 (no conservation interest): River stretches belonging to watersheds where G. aculeatus presence was not confirmed. This level was also assigned to river stretches with absence of the species and with an occurrence probability of less than 20% but belonging to watersheds in which, in other river stretches, the presence of the species has already been confirmed.
- Level 1 (moderate importance for conservation): River stretches with the absence of the species and where the probability of occurrence ranges between 20% and 40% but belonging to watersheds that cover other river stretches in which the presence of the species has already been confirmed.
- Level 2 (high importance for conservation): River stretches with the absence of the species and where the probability of occurrence is above to 40% but belonging to watersheds that cover other river stretches in which the presence of the species has already been confirmed.
- Level 3 (maximum importance for conservation): River stretches with confirmed presence of G. aculeatus and stretches upstream and downstream of the presence with a probability of occurrence greater than 10%.
3. Results
3.1. Distribution of G. aculeatus
3.2. Relationship between G. aculeatus Occurrence and Environmental Predictors
3.3. Spatial Predictions of G. aculeatus Probability of Occurrence
3.4. Map of Conservation Priorities
4. Discussion
4.1. Influence of Environmental Variables on the Distribution of G. aculeatus
4.2. Prioritizing Conservation of G. aculeatus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Bell, M.A.; Foster, S.A. The Evolutionary Biology of the Threespine Stickleback; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Cabral, M.J.; Almeida, J.; Almeida, P.R.; Dellinger, T.; Ferrand de Almeida, N.; Oliveira, M.E.; Palmeirim, J.M.; Queiroz, A.M.; Rogado, L.; Santos Reis, M. (Eds.) Livro Vermelho Dos Vertebrados De Portugal: Peixes Dulciaquıícolas E Migradores, Anfíbios, Repteis, Aves E Mamíferos; Instituto da Conservação da Natureza: Lisboa, Portugal, 2005; 660p. [Google Scholar]
- Hagen, D.W. Isolating Mechanisms in Threespine Sticklebacks (Gasterosteus). J. Fish. Res. Board Can. 1967, 24, 1637–1692. [Google Scholar] [CrossRef]
- Berra, T.M. Freshwater Fish Distribution; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Crivelli, A.J.; Britton, R.H. Life history adaptations of Gasterosteus aculeatus in a Mediterranean wetland. Environ. Biol. Fishes 1987, 18, 109–125. [Google Scholar] [CrossRef]
- Rind, K.; Rodriguez-Barucg, Q.; Nicolas, D.; Cucchi, P.; Lignot, J. Morphological and physiological traits of Mediterranean sticklebacks living in the Camargue wetland (Rhone river delta). J. Fish Biol. 2020, 97, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.A.; Baker, J.A.; Bell, B.A. The case for conserving threespine stickleback populations: Protecting an adaptive radiation. Fisheries 2003, 28, 10–18. [Google Scholar] [CrossRef]
- Sanz, N.; Araguas, R.M.; Vidal, O.; Viñas, J. Glacial refuges for three-spined stickleback in the Iberian Peninsula: Mitochondrial DNA phylogeography. Freshw. Biol. 2015, 60, 1794–1809. [Google Scholar] [CrossRef]
- Alexandre, C.M.; Almeida, P.R. Summer survival and habitat characteristics of a threespine stickleback (Gasterosteus aculeatus L.) Southern European Population. Limnetica 2009, 28, 125–138. [Google Scholar] [CrossRef]
- Clavero, M.; Pou-Rovira, Q.; Zamora, L. Biology and habitat use of three-spined stickleback (Gasterosteus aculeatus) in intermittent Mediterranean streams. Ecol. Freshw. Fish 2009, 18, 550–559. [Google Scholar] [CrossRef]
- Normand, S.; Treier, U.A.; Randin, C.; Vittoz, P.; Guisan, A.; Svenning, J.-C. Importance of abiotic stress as a range-limit determinant for European plants: Insights from species responses to climatic gradients. Glob. Ecol. Biogeogr. 2009, 18, 437–449. [Google Scholar] [CrossRef]
- Araguas, R.M.; Vidal, O.; Pla, C.; Sanz, N. High genetic diversity of the endangered Iberian three-spined stickleback (Gasterosteus aculeatus) at the Mediterranean edge of its range. Freshw. Biol. 2011, 57, 143–154. [Google Scholar] [CrossRef]
- Mayden, R.L.; Page, L.M.; Burr, B.M. A Field Guide to Freshwater Fishes of North America North of Mexico. Copeia 1992, 1992, 920. [Google Scholar] [CrossRef]
- Gorman, O.T.; Karr, J.R. Habitat Structure and Stream Fish Communities. Ecology 1978, 59, 507–515. [Google Scholar] [CrossRef]
- Lanka, R.P.; Hubert, W.A.; Wesche, T.A. Relations of Geomorphology to Stream Habitat and Trout Standing Stock in Small Rocky Mountain Streams. Trans. Am. Fish. Soc. 1987, 116, 21–28. [Google Scholar] [CrossRef]
- Ferreira, A.; Quintella, B.; Maia, C.; Mateus, C.; Alexandre, C.; Capinha, C.; Almeida, P. Influence of macrohabitat preferences on the distribution of European brook and river lampreys: Implications for conservation and management. Biol. Conserv. 2013, 159, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Rieman, B.E.; McIntyre, J.D. Occurrence of Bull Trout in Naturally Fragmented Habitat Patches of Varied Size. Trans. Am. Fish. Soc. 1995, 124, 285–296. [Google Scholar] [CrossRef]
- Watson, G.; Hillman, T.W. Factors Affecting the Distribution and Abundance of Bull Trout: An Investigation at Hierarchical Scales. North Am. J. Fish. Manag. 1997, 17, 237–252. [Google Scholar] [CrossRef]
- Porter, M.S.; Rosenfeld, J.; Parkinson, E.A. Predictive Models of Fish Species Distribution in the Blackwater Drainage, British Columbia. North Am. J. Fish. Manag. 2000, 20, 349–359. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Diniz-Filho, J.A.F.; Bini, L.M.; Rangel, T.F.; Loyola, R.D.; Hof, C.; Noguã©S-Bravo, D.; AraãºJo, M.B. Partitioning and mapping uncertainties in ensembles of forecasts of species turnover under climate change. Ecography 2009, 32, 897–906. [Google Scholar] [CrossRef]
- Zhang, X.; Vincent, A.C. Predicting distributions, habitat preferences and associated conservation implications for a genus of rare fishes, seahorses (Hippocampus spp.). Divers. Distrib. 2018, 24, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Gasith, A.; Resh, V.H. Streams in Mediterranean Climate Regions: Abiotic Influences and Biotic Responses to Predictable Seasonal Events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- ICA. Iberian Climate Atlas: Air Temperature and Precipitation (1971–2000); Department of the State Meteorological Agency of Spain, and Portugal, Department of Meteorology and Climatology of the Institute of Meteorology: Madrid, Spain, 2011; 80p. [Google Scholar]
- IPMA–Instituto Português do Mar e da Atmosfera. Área Educativa–Clima de Portugal Continental. 2020. Available online: www.portal.pt/pt/educativa/tempo.clima/ (accessed on 16 October 2020).
- Central Intelligence Agency, Portugal: The world factbook. Langley, Virginia: Central Intelligence Agency. 2010. Available online: www.cia.gov/library/publications/the-world-factbook/geos/po.html (accessed on 15 October 2020).
- Inag, I.P. Manual Para A Avaliação Biológica da Qualidade da Água em Sistemas Fluviais Segundo A Directiva Quadro da Água Protocolo de Amostragem E Análise Para A Fauna Piscícola; Ministério do Ambiente, do Ordenamento do Território e do Desenvolvimento Regional. Instituto da Água, I.P., Instituto da água: Lisbon, Portugal, 2008. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- ESRI. ArcGIS/ArcInfo for Windows XP. Version 9.3.1; Environmental System Research Institute: Redlands, CA, USA, 2009. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Fernández, C.; San Miguel, E.; Amaro, R.; Hermida, M. Espinoso–Gasterosteus aculeatus. In Enciclopedia Virtual De Los Vertebrados Españoles (Virtual Encyclopaedia of Spanish vertebrates); Carrascal, L.M., Salvador, A., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2006. [Google Scholar]
- FAO/IIASA/ISRIC/ISSCAS/JRC. Harmonized World Soil Database. Version 1.1. FAO: Rome, Italy; IIASA: Laxenburg, Austria, 2009. [Google Scholar]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The Natural Flow Regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Zeiringer, B.; Seliger, C.; Greimel, F.; Schmutz, S. River Hydrology, Flow Alteration, and Environmental Flow. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer: Cham, Switzerland, 2018; Volume 8. [Google Scholar]
- SAGA. System for Automated Geoscientific Analysis. Version 2.0.6; SAGA User Group Association: Boston, MA, USA, 2010. [Google Scholar]
- Böhner, J.; Köthe, R.; Conrad, O.; Gross, J.; Ringeler, A.; Selige, T. Soil regionalisation by means of terrain analysis and process parameterization. In Soil Classification 2001; Micheli, E., Nachtergaele, F., Montanarella, L., Eds.; The European Soil Bureau, Joint Research Centre: Ispra, Italy, 2002; pp. 213–222. [Google Scholar]
- Moore, I.D.; Burch, G.J.; MacKenzie, D.H. Topographic Effects on the Distribution of Surface Soil Water and the Location of Ephemeral Gullies. Am. Soc. Agric. Biol. Eng. 1988, 31, 1098–1107. [Google Scholar] [CrossRef]
- Erickson, T.R.; Stefan, H.G. Linear Air/Water Temperature Correlations for Streams during Open Water Periods. J. Hydrol. Eng. 2000, 5, 317–321. [Google Scholar] [CrossRef]
- Lassalle, G.; Béguer, M.; Beaulaton, L.; Rochard, E. Diadromous fish conservation plans need to consider global warming issues: An approach using biogeographical models. Biol. Conserv. 2008, 141, 1105–1118. [Google Scholar] [CrossRef]
- Amoatey, P.; Baawain, M.S. Effects of pollution on freshwater aquatic organisms. Water Environ. Res. 2019, 91, 1272–1287. [Google Scholar] [CrossRef] [Green Version]
- Kroll, S.A.; Llacer, C.N.; Cano, M.D.L.C.; Heras, J.D.L. The inuence of land use on water quality and macroinvertebrate biotic indices in rivers within Castilla-La Mancha (Spain). Limnetica 2009, 28, 203–214. [Google Scholar] [CrossRef]
- EEA. Corine Land Cover 2000. Version 13. European Environmental Agency. 2010. Available online: www.eea.europa.eu/data-and-maps/data/clc-2000-raster (accessed on 17 March 2020).
- LandScan, Geographic Information Science and Technology. OakRidge National Laboratory. UT-Battelle. US Department of Energy. 2010. Available online: www.landscan.ornl.gov/ (accessed on 24 March 2020).
- Naimi, B.; Araújo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef] [Green Version]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning with Applications in R; Springer: New York, NY, USA, 2017. [Google Scholar]
- Graham, M.H. Confronting Multicollinearity in Ecological Multiple Regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Boavida-Portugal, J.; Rosa, R.; Calado, R.; Pinto, M.; Boavida-Portugal, I.; Araújo, M.B.; Guilhaumon, F. Climate change impacts on the distribution of coastal lobsters. Mar. Biol. 2018, 165, 186. [Google Scholar] [CrossRef]
- Crossman, N.D.; Bass, D.A. Application of common predictive habitat techniques for post-border weed risk management. Divers. Distrib. 2007, 14, 213–224. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Ray, S.; Saikia, S.K. Indicator environmental variables in regulating the distribution patterns of small freshwater fish Amblypharyngodon mola in India and Bangladesh. Ecol. Indic. 2020, 120, 106906. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Rueda, M.; Rodríguez, M. What Do Range Maps and Surveys Tell Us About Diversity Patterns? Folia Geobot. 2008, 43, 345–355. [Google Scholar] [CrossRef]
- Arboleya, E.; Fernández, S.; Clusa, L.; Dopico, E.; Garcia-Vazquez, E. River Connectivity is Crucial for Safeguarding Biodiversity but May be Socially Overlooked. Insights From Spanish University Students. Front. Environ. Sci. 2021, 9. [Google Scholar] [CrossRef]
- Apaydin, H.; Anli, A.S.; Ozturk, F. Evaluation of topographical and geographical effects on some climatic parameters in the Central Anatolia Region of Turkey. Int. J. Clim. 2010, 31, 1264–1279. [Google Scholar] [CrossRef]
- Camana, M.; Dala-Corte, R.B.; Becker, F.G. Relation between species richness and stream slope in riffle fish assemblages is dependent on spatial scale. Environ. Biol. Fishes 2016, 99, 603–612. [Google Scholar] [CrossRef]
- Cluis, D.A. Relationship between Stream Water Temperature and Ambient Air Temperature. Water Policy 1972, 3, 65–71. [Google Scholar] [CrossRef]
- Baggerman, B. An Experimental Study on the Timing of Breeding and Migration in the Three-Spined Stickleback. Arch. Néerlandaises De Zool. 1958, 12, 105–317. [Google Scholar] [CrossRef] [Green Version]
- Borg, B.; Van Veen, T. Seasonal effects of photoperiod and temperature on the ovary of the three-spined stickleback, Gasterosteus aculeatus L. Can. J. Zool. 1982, 60, 3387–3393. [Google Scholar] [CrossRef]
- Dubeux, J.C.B.; Sollenberger, L.E. Nutrient cycling in grazed pastures. Manag. Strateg. Sustain. Cattle Prod. South. Pastures 2020, 1, 59–75. [Google Scholar] [CrossRef]
- Oberdoff, T.; Guegan, J.-F.; Hugueny, B. Global scale patterns of fish species richness in rivers. Ecography 1995, 18, 345–352. [Google Scholar] [CrossRef]
- Cohen, S.; Wan, T.; Islam, T.; Syvitski, J. Global river slope: A new geospatial dataset and global-scale analysis. J. Hydrol. 2018, 563, 1057–1067. [Google Scholar] [CrossRef]
- Shumie, M.C. River Slope and Roughness Impact on Downstream Hydraulic Structures. J. Earth Sci. Clim. Chang. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Bonada, N.; Resh, V.H. Mediterranean-climate streams and rivers: Geographically separated but ecologically comparable freshwater systems. Hydrobiologia 2013, 719, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Harvey, B.C. Susceptibility of Young-of-the-Year Fishes to Downstream Displacement by Flooding. Trans. Am. Fish. Soc. 1987, 116, 851–855. [Google Scholar] [CrossRef]
- Chun, S.N.; Cocherell, S.A.; Cocherell, D.E.; Miranda, J.B.; Jones, G.J.; Graham, J.; Cech, J.J., Jr. Displacement, velocity preference, and substrate use of three native California stream fishes in simulated pulsed flows. Environ. Biol. Fishes 2011, 90, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Taylor, E.B.; McPhail, J.D. Prolonged and burst swimming in anadromous and freshwater threespine stickleback, Gasterosteus aculeatus. Can. J. Zool. 1986, 64, 416–420. [Google Scholar] [CrossRef]
- Karve, A.D.; von Hippel, F.A.; Bell, M.A. Isolation between sympatric anadromous and resident threespine stickleback species in Mud Lake, Alaska. Environ. Biol. Fishes 2007, 81, 287–296. [Google Scholar] [CrossRef]
- Arai, T.; Ueno, D.; Kitamura, T.; Goto, A. Habitat preference and diverse migration in threespine sticklebacks, Gasterosteus aculeatus and G. nipponicus. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Raeymaekers, J.A.M.; Maes, G.E.; Audenaert, E.; Volckaert, F.A.M. Detecting Holocene divergence in the anadromous-freshwater three-spined stickleback (Gasterosteus aculeatus) system. Mol. Ecol. 2005, 14, 1001–1014. [Google Scholar] [CrossRef]
- Mota, M.; Antunes, C. First report on the status of Allis shad (Alosa alosa) in the Minho River (Northwestern Iberian Peninsula). J. Appl. Ichthyol. 2011, 27, 56–59. [Google Scholar] [CrossRef]
- Almeida, P.R.; Quintella, B.R.; Mateus, C.S.; Alexandre, C.M.; Pedro, S. Diadromous fish in Portugal: Status, threats and management guidelines. In Sustainable Development of the Ocean: A Necessity; Bebiano, M.J., Guerreiro, J., Carvalho, T., Gameiro, M.I., Eds.; Universidade do Algarve editora: Algarve, Portugal, 2018; pp. 189–213. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Code | Range |
|---|---|---|
| Geomorphology | ||
| Altitude (m) | altitude | 0–1959 |
| Slope (°) | slope | 0–48.689 |
| Distance to coast (m) | dist_coast | 0–357,420 |
| Silt (%) | silt | 3.6–20.3 |
| Sand (%) | sand | 19.1–85.9 |
| Climate | ||
| Annual mean temperature (°C) | tempmean | 6.6–17.8 |
| Maximum temperature of warmest month (°C) | tempmax | 20.4–33.8 |
| Annual precipitation (mm) | precip | 477–1880 |
| Precipitation of driest month (mm) | precipdriest | 1–35 |
| Hydrology | ||
| Flow accumulation (no. of cells) | f_accum | 0–9,764,683 |
| Flow weight with rainfall (no. of cells) | f_weight_rain | 0–267,597,088 |
| WTI (Wetness Topographic Index) | wti | 0–20.07 |
| SPI (Stream Power Index) | spi | −3.5 × 109 to 4.5 × 108 |
| Environmental stressors | ||
| Artificial surfaces (no. of cells) | use_art | 0–6083 |
| Agricultural areas (no. of cells) | use_agr | 0–218,888 |
| Forest and semi-natural areas (no. of cells) | use_forest | 0–161,346 |
| Population (n/km²) | populat | 0–15,304 |
| Catchment | River | River Stretch (km) | Maintenance of Connectivity with Sea |
|---|---|---|---|
| Mira | Pilriteiro Stream | 15 | Yes |
| Sado | Gema Stream | 17 | No |
| Grândola Stream | 21 | No | |
| Marateca Stream | 8 | Yes | |
| Tejo | Almansor River | 10 | Yes |
| Tejo River | 57 | Yes | |
| Almonda River | 18 | No | |
| Zêzere River | 7 | Yes | |
| Pisão Stream | 5 | No | |
| Lis | Lis River | 15 | Yes |
| Leça Stream | 15 | Yes | |
| Mondego | Arunca River | 29 | Yes |
| Anços River | 11 | Yes | |
| Ança Stream | 27 | Yes | |
| Vouga | Vale da Corujeira Stream | 31 | Yes |
| Varziela Stream | 13 | Yes | |
| Palhal Stream | 15 | Yes | |
| Vouga River | 36 | Yes | |
| Águeda River | 14 | Yes | |
| Cértima River | 30 | Yes | |
| Negra Stream | 13 | Yes | |
| Seixo Stream | 3 | Yes | |
| Cáster Stream | 5 | Yes | |
| Cávado | Milhases Stream | 5 | Yes |
| Lima | Lima River | 32 | Yes |
| Vez River | 13 | Yes | |
| Estorãos River | 8 | Yes | |
| Âncora | Âncora River | 8 | Yes |
| Minho | Minho River | 48 | Yes |
| Coura River | 11 | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, A.; Boavida-Portugal, J.; Almeida, P.R.; Silva, S.; Alexandre, C.M. Macro-Habitat Suitability for Threespine Stickleback (Gasterosteus aculeatus L.) Near the Southern Limit of Its Global Distribution: Implications for Species Management and Conservation. Fishes 2022, 7, 271. https://doi.org/10.3390/fishes7050271
Moreira A, Boavida-Portugal J, Almeida PR, Silva S, Alexandre CM. Macro-Habitat Suitability for Threespine Stickleback (Gasterosteus aculeatus L.) Near the Southern Limit of Its Global Distribution: Implications for Species Management and Conservation. Fishes. 2022; 7(5):271. https://doi.org/10.3390/fishes7050271
Chicago/Turabian StyleMoreira, Andre, Joana Boavida-Portugal, Pedro R. Almeida, Sara Silva, and Carlos M. Alexandre. 2022. "Macro-Habitat Suitability for Threespine Stickleback (Gasterosteus aculeatus L.) Near the Southern Limit of Its Global Distribution: Implications for Species Management and Conservation" Fishes 7, no. 5: 271. https://doi.org/10.3390/fishes7050271