
DNA Methylation and Detoxification in the Earthworm Lumbricus terrestris Exposed to Cadmium and the DNA Demethylation Agent 5-aza-2′-deoxycytidine



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup and Sampling
2.2. RNA Extraction and cDNA Synthesis
2.3. Quantitative Real-Time PCR
2.4. Protein Extraction and DNMT Activity
2.5. Quantification of 5-Methylcytosine (5mC) and 5-Hydroxymethylcytosine (5hmC)
2.6. Statistical Analysis
3. Results
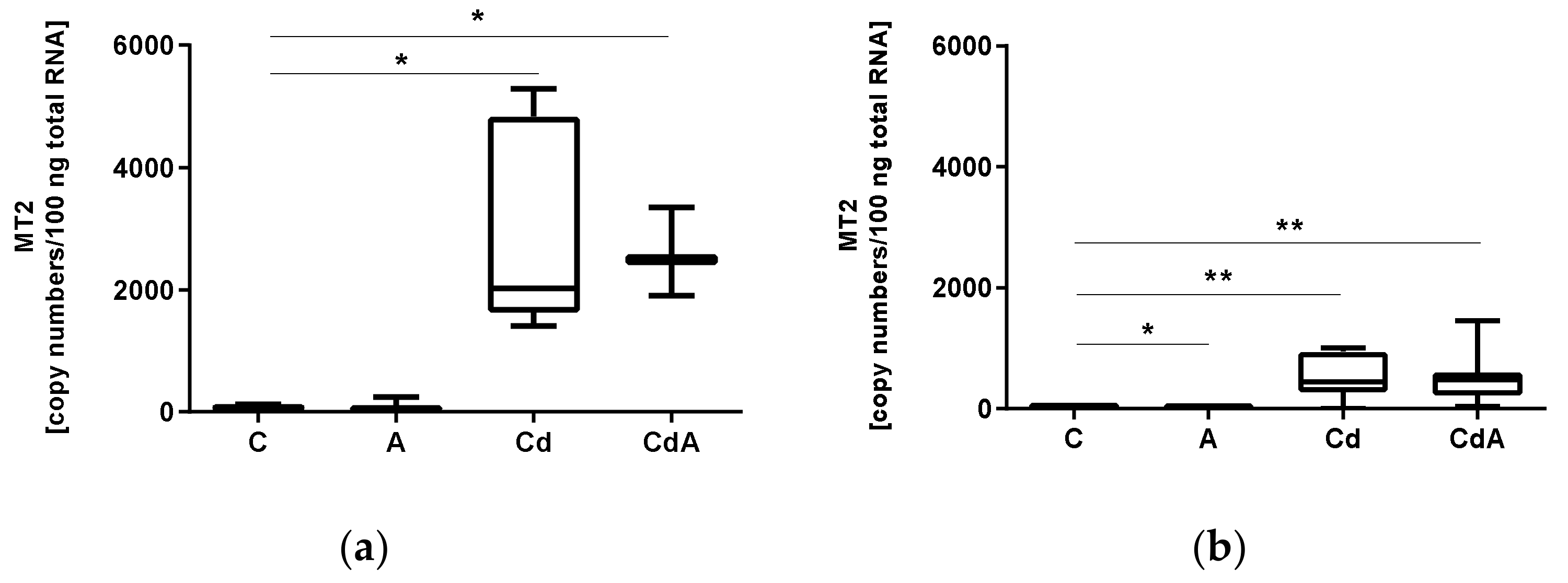
3.1. Quantification of Gene Expression
3.2. Global Methylation Level and DNMT Activity
3.3. Global 5hmc
3.4. Multidimensional Scaling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lemtiri, A.; Colinet, G.; Alabi, T.; Cluzeau, D.; Zirbes, L.; Haubruge, É.; Francis, F. Impacts of Earthworms on Soil Components and Dynamics. A Review. 2014. Available online: https://popups.uliege.be/1780-4507/index.php?id=10881 (accessed on 18 June 2021).
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. Int. Sch. Res. Not. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Asami, T. Pollution of Soils by Cadmium. In Changing Metal Cycles and Human Health; Springer: Berlin/Heidelberg, Germany, 1984; pp. 95–111. [Google Scholar]
- Bridges, C.C.; Zalups, R.K. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [Green Version]
- Duncan, K. Metallothioneins and Related Chelators. Metal Ions in Life Sciences Vol. 5. Edited by Astrid Sigel, Helmut Sigel and Roland, K.O. Sigel. Angew. Chem. Int. Ed. 2009, 48, 7966–7967. [Google Scholar] [CrossRef]
- Ma, Y.; Rivera-Ingraham, G.A.; Nommick, A.; Bickmeyer, U.; Roeder, T. Copper and cadmium administration induce toxicity and oxidative stress in the marine flatworm Macrostomum lignano. Aquat. Toxicol. 2020, 221, 105428. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Pedrini-Martha, V.; Schnegg, R.; Schäfer, G.G.; Lieb, B.; Salvenmoser, W.; Dallinger, R. Responsiveness of metallothionein and hemocyanin genes to cadmium and copper exposure in the garden snail Cornu aspersum. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2020, 335, 228–238. [Google Scholar] [CrossRef]
- Tang, S.; Allagadda, V.; Chibli, H.; Nadeau, J.L.; Mayer, G.D. Comparison of cytotoxicity and expression of metal regulatory genes in zebrafish (Danio rerio) liver cells exposed to cadmium sulfate, zinc sulfate and quantum dots. Metallomics 2013, 5, 1411–1422. [Google Scholar] [CrossRef]
- Probst, G.S.; Bousquet, W.F.; Miya, T.S. Correlation of hepatic metallothionein concentrations with acute cadmium toxicity in the mouse. Toxicol. Appl. Pharmacol. 1977, 39, 61–69. [Google Scholar] [CrossRef]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Flores, K.B.; Wolschin, F.; Amdam, G.V. The Role of Methylation of DNA in Environmental Adaptation. Integr. Comp. Biol. 2013, 53, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Zamudio, R.; Ha, H.C. Environmental epigenetics in metal exposure. Epigenetics 2011, 6, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, Mechanisms and Clinical Perspective. Semin. Reprod. Med. 2009, 27, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trijau, M.; Asselman, J.; Armant, O.; Adam-Guillermin, C.; De Schamphelaere, K.A.C.; Alonzo, F. Transgenerational DNA Methylation Changes in Daphnia magna Exposed to Chronic γ Irradiation. Environ. Sci. Technol. 2018, 52, 4331–4339. [Google Scholar] [CrossRef] [PubMed]
- Mirbahai, L.; Chipman, J.K. Epigenetic memory of environmental organisms: A reflection of lifetime stressor exposures. Mutat. Res. Toxicol. Environ. Mutagen. 2014, 764–765, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Jeremias, G.; Barbosa, J.; Marques, S.M.; De Schamphelaere, K.A.; Van Nieuwerburgh, F.; Deforce, D.; Gonçalves, F.J.; Pereira, J.L.; Asselman, J. Transgenerational Inheritance of DNA Hypomethylation in Daphnia magna in Response to Salinity Stress. Environ. Sci. Technol. 2018, 52, 10114–10123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šrut, M. Ecotoxicological epigenetics in invertebrates: Emerging tool for the evaluation of present and past pollution burden. Chemosphere 2021, 282, 131026. [Google Scholar] [CrossRef]
- Holliday, R. Epigenetics: A Historical Overview. Epigenetics 2006, 1, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Riggs, A.D. X inactivation, differentiation, and DNA methylation. Cytogenet Genome Res. 1975, 14, 9–25. [Google Scholar] [CrossRef]
- Holliday, R.; Pugh, J.E. DNA Modification Mechanisms and Gene Activity During Development. Science 1975, 187, 226–232. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for De Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Lyko, F. The DNA methyltransferase family: A versatile toolkit for epigenetic regulation. Nat. Rev. Genet. 2017, 19, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Sassa, A.; Kanemaru, Y.; Kamoshita, N.; Honma, M.; Yasui, M. Mutagenic consequences of cytosine alterations site-specifically embedded in the human genome. Genes Environ. 2016, 38, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagiwada, S.; Kurimoto, K.; Hirota, T.; Yamaji, M.; Saitou, M. Replication-coupled passive DNA demethylation for the erasure of genome imprints in mice. EMBO J. 2012, 32, 340–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Dahlet, T.; Lleida, A.A.; Al Adhami, H.; Dumas, M.; Bender, A.; Ngondo, R.P.; Tanguy, M.; Vallet, J.; Auclair, G.; Bardet, A.F.; et al. Genome-wide analysis in the mouse embryo reveals the importance of DNA methylation for transcription integrity. Nat. Commun. 2020, 11, 3153. [Google Scholar] [CrossRef]
- Héberlé, É.; Bardet, A.F. Sensitivity of transcription factors to DNA methylation. Essays Biochem. 2019, 63, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Kulis, M.; Esteller, M. DNA Methylation and Cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar] [CrossRef]
- Seelan, R.S.; Mukhopadhyay, P.; Pisano, M.M.; Greene, R.M. Effects of 5-aza-2′-deoxycytidine (decitabine) on gene expression. Drug Metab. Rev. 2017, 50, 193–207. [Google Scholar] [CrossRef]
- Filì, C.; Candoni, A.; Zannier, M.E.; Olivieri, J.; Imbergamo, S.; Caizzi, M.; Nadali, G.; Di Bona, E.; Ermacora, A.; Gottardi, M.; et al. Efficacy and toxicity of Decitabine in patients with acute myeloid leukemia (AML): A multicenter real-world experience. Leuk. Res. 2019, 76, 33–38. [Google Scholar] [CrossRef]
- Kumar, S.; Kim, Y. An endoparasitoid wasp influences host DNA methylation. Sci. Rep. 2017, 7, 43287. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, J.; Lyu, H.; Song, Q.; Feng, Q.; Xiang, H.; Zheng, S. DNA methylation mediates BmDeaf1-regulated tissue- and stage-specific expression of BmCHSA-2b in the silkworm, Bombyx mori. Epigenet. Chromatin 2018, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planques, A.; Kerner, P.; Ferry, L.; Grunau, C.; Gazave, E.; Vervoort, M. DNA methylation atlas and machinery in the developing and regenerating annelid Platynereis dumerilii. BMC Biol. 2021, 19, 148. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, J.; Momparler, R.L. Incorporation of 5-aza-2′-deoxycytidine-5′-triphosphate into DNA. Interactions with mammalian DNA polymerase alpha and DNA methylase. Mol. Pharmacol. 1983, 24, 109–114. [Google Scholar]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [Green Version]
- Momparler, R.L. Pharmacology of 5-aza-2′-deoxycytidine (decitabine). Semin. Hematol. 2005, 42, S9–S16. [Google Scholar] [CrossRef]
- Creusot, F.; Acs, G.; Christman, J.K. Inhibition of DNA methyltransferase and induction of Friend erythroleukemia cell differentiation by 5-azacytidine and 5-aza-2′-deoxycytidine. J. Biol. Chem. 1982, 257, 2041–2048. [Google Scholar] [CrossRef]
- Taylor, S.M.; Jones, P.A. Mechanism of action of eukaryotic DNA methyltransferase: Use of 5-azacytosine-containing DNA. J. Mol. Biol. 1982, 162, 679–692. [Google Scholar] [CrossRef]
- Akcha, F.; Barranger, A.; Bachère, E. Genotoxic and epigenetic effects of diuron in the Pacific oyster: In vitro evidence of interaction between DNA damage and DNA methylation. Environ. Sci. Pollut. Res. 2020, 28, 8266–8280. [Google Scholar] [CrossRef]
- Bachère, E.; Barranger, A.; Bruno, R.; Rouxel, J.; Menard, D.; Piquemal, D.; Akcha, F. Parental diuron-exposure alters offspring transcriptome and fitness in Pacific oyster Crassostrea gigas. Ecotoxicol. Environ. Saf. 2017, 142, 51–58. [Google Scholar] [CrossRef]
- Gatzmann, F.; Falckenhayn, C.; Gutekunst, J.; Hanna, K.; Raddatz, G.; Carneiro, V.C.; Lyko, F. The methylome of the marbled crayfish links gene body methylation to stable expression of poorly accessible genes. Epigenet. Chromatin 2018, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Šrut, M.; Drechsel, V.; Höckner, M. Low levels of Cd induce persisting epigenetic modifications and acclimation mechanisms in the earthworm Lumbricus terrestris. PLoS ONE 2017, 12, e0176047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homa, J.; Klimek, M.; Kruk, J.; Cocquerelle, C.; Vandenbulcke, F.; Plytycz, B. Metal-specific effects on metallothionein gene induction and riboflavin content in coelomocytes of Allolobophora chlorotica. Ecotoxicol. Environ. Saf. 2010, 73, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Aigner, G.P.; Pittl, V.; Fiechtner, B.; Egger, B.; Šrut, M.; Höckner, M. Common mechanisms cannot explain time- and dose-dependent DNA methylation changes in earthworms exposed to cadmium. Sci. Total Environ. 2021, 812, 151468. [Google Scholar] [CrossRef] [PubMed]
- Jeremias, G.; Gonçalves, F.J.M.; Pereira, J.L.; Asselman, J. Prospects for incorporation of epigenetic biomarkers in human health and environmental risk assessment of chemicals. Biol. Rev. 2020, 95, 822–846. [Google Scholar] [CrossRef] [Green Version]
- Vandegehuchte, M.B.; Janssen, C.R. Epigenetics in an ecotoxicological context. Mutat. Res. Toxicol. Environ. Mutagen. 2014, 764–765, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Drechsel, V.; Schauer, K.; Šrut, M.; Höckner, M. Regulatory Plasticity of Earthworm wMT-2 Gene Expression. Int. J. Mol. Sci. 2017, 18, 1113. [Google Scholar] [CrossRef] [Green Version]
- Du Preez, P.H.; Breeds, K.; Burger, N.F.V.; Swiegers, H.W.; Truter, J.C.; Botha, A.-M. DNA Methylation and Demethylation Are Regulated by Functional DNA Methyltransferases and DnTET Enzymes in Diuraphis noxia. Front. Genet. 2020, 11, 452. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 26 December 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 26 December 2021).
- Kang, S.W.; Madkour, M.; Kuenzel, W.J. Tissue-Specific Expression of DNA Methyltransferases Involved in Early-Life Nutritional Stress of Chicken, Gallus gallus. Front. Genet. 2017, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Zupkovitz, G.; Kabiljo, J.; Kothmayer, M.; Schlick, K.; Schöfer, C.; Lagger, S.; Pusch, O. Analysis of Methylation Dynamics Reveals a Tissue-Specific, Age-Dependent Decline in 5-Methylcytosine Within the Genome of the Vertebrate Aging Model Nothobranchius furzeri. Front. Mol. Biosci. 2021, 8, 627143. [Google Scholar] [CrossRef]
- Waalkes, M.P.; Miller, M.S.; Wilson, M.J.; Bare, R.M.; McDowell, A.E. Increased metallothionein gene expression in 5-aza-2′-deoxycytidine-induced resistance to cadmium cytotoxicity. Chem. Interact. 1988, 66, 189–204. [Google Scholar] [CrossRef]
- Amarasinghe, H.E.; Clayton, C.I.; Mallon, E.B. Methylation and worker reproduction in the bumblebee (Bombus terrestris). Proc. R. Soc. B Boil. Sci. 2014, 281, 20132502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homa, J.; Bzowska, M.; Klimek, M.; Plytycz, B. Flow cytometric quantification of proliferating coelomocytes non-invasively retrieved from the earthworm, Dendrobaena veneta. Dev. Comp. Immunol. 2008, 32, 9–14. [Google Scholar] [CrossRef]
- Cook, N.; Parker, D.J.; Tauber, E.; Pannebakker, B.A.; Shuker, D.M.; Dj, P.; Ba, P. Validating the Demethylating Effects of 5-aza-2′-deoxycytidine in Insects Requires a Whole-Genome Approach. Am. Nat. 2019, 194, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Tian, X.; Yang, J.; Cui, J.; Jiang, S.; Shi, R.; Liu, Y.; Liu, X.; Xu, W.; Xie, W.; et al. 5-aza-2′-deoxycytidine, a DNA methylation inhibitor, induces cytotoxicity, cell cycle dynamics and alters expression of DNA methyltransferase 1 and 3A in mouse hippocampus-derived neuronal HT22 cells. J. Toxicol. Environ. Health Part A 2017, 80, 1222–1229. [Google Scholar] [CrossRef]
- Honeywell, R.J.; Sarkisjan, D.; Kristensen, M.H.; de Klerk, D.J.; Peters, G.J. DNA methyltransferases expression in normal tissues and various human cancer cell lines, xenografts and tumors. Nucleosides Nucleotides Nucleic Acids 2018, 37, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Honeywell, R.J.; Sarkisjan, D.; Kathmann, I.; Kristensen, M.H.; Peters, G.J. Sensitive liquid chromatography mass spectrometry (LC-MS) assay reveals novel insights on DNA methylation and incorporation of gemcitabine, its metabolite difluorodeoxyuridine, deoxyuridine, and RX-3117 into DNA. Nucleosides Nucleotides Nucleic Acids 2016, 35, 652–662. [Google Scholar] [CrossRef]
- Chaney, R.L. Cadmium and Zinc. In Trace Elements in Soils; John Wiley & Sons, Ltd.: Chichester, UK, 2010; pp. 409–439. [Google Scholar]
- Das, P.; Samantaray, S.; Rout, G.R. Studies on cadmium toxicity in plants: A review. Environ. Pollut. 1997, 98, 29–36. [Google Scholar] [CrossRef]
- Tang, L.; Qiu, R.; Tang, Y.; Wang, S. Cadmium–zinc exchange and their binary relationship in the structure of Zn-related proteins: A mini review. Metallomics 2014, 6, 1313–1323. [Google Scholar] [CrossRef]
- He, S.; Wang, F.; Yang, L.; Guo, C.; Wan, R.; Ke, A.; Xu, L.; Hu, G.; Xu, X.; Shen, J.; et al. Expression of DNMT1 and DNMT3a Are Regulated by GLI1 in Human Pancreatic Cancer. PLoS ONE 2011, 6, e27684. [Google Scholar] [CrossRef] [Green Version]
- Kishikawa, S.; Murata, T.; Kimura, H.; Shiota, K.; Yokoyama, K.K. Regulation of transcription of the Dnmt1 gene by Sp1 and Sp3 zinc finger proteins. JBIC J. Biol. Inorg. Chem. 2002, 269, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Hodge, D.R.; Xiao, W.; Clausen, P.A.; Heidecker, G.; Szyf, M.; Farrar, W.L. Interleukin-6 Regulation of the Human DNA Methyltransferase (HDNMT) Gene in Human Erythroleukemia Cells. J. Biol. Chem. 2001, 276, 39508–39511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, M.; Khan, N.M.; Ahmad, N.; Green, J.; Novak, K.; Haqqi, T.M. Genetic Inactivation of ZCCHC 6 Suppresses Interleukin-6 Expression and Reduces the Severity of Experimental Osteoarthritis in Mice. Arthritis Rheumatol. 2019, 71, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Pestana, J.L.; Novais, S.C.; Norouzitallab, P.; Vandegehuchte, M.B.; Bossier, P.; De Schamphelaere, K.A. Non-lethal heat shock increases tolerance to metal exposure in brine shrimp. Environ. Res. 2016, 151, 663–670. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Treatment | Statistical Test | p Value |
|---|---|---|---|
| MT2 | C | Student’s t-test | 0.0117 |
| A | Mann–Whitney U test | 0.0606 | |
| Cd | Welch’s test | 0.0324 | |
| CdA | Student’s t-test | <0.0001 | |
| DNMT1 | C | Welch’s test | 0.0216 |
| A | Welch’s test | 0.019 | |
| Cd | Mann–Whitney U test | 0.0455 | |
| CdA | Welch’s test | 0.2101 | |
| TET | C | Welch’s test | 0.0025 |
| A | Student’s t-test | 0.0208 | |
| Cd | Welch’s test | 0.0025 | |
| CdA | Student’s t-test | 0.0008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aigner, G.P.; Nenning, P.; Fiechtner, B.; Šrut, M.; Höckner, M. DNA Methylation and Detoxification in the Earthworm Lumbricus terrestris Exposed to Cadmium and the DNA Demethylation Agent 5-aza-2′-deoxycytidine. Toxics 2022, 10, 100. https://doi.org/10.3390/toxics10020100
Aigner GP, Nenning P, Fiechtner B, Šrut M, Höckner M. DNA Methylation and Detoxification in the Earthworm Lumbricus terrestris Exposed to Cadmium and the DNA Demethylation Agent 5-aza-2′-deoxycytidine. Toxics. 2022; 10(2):100. https://doi.org/10.3390/toxics10020100
Chicago/Turabian StyleAigner, Gerhard P., Pamela Nenning, Birgit Fiechtner, Maja Šrut, and Martina Höckner. 2022. "DNA Methylation and Detoxification in the Earthworm Lumbricus terrestris Exposed to Cadmium and the DNA Demethylation Agent 5-aza-2′-deoxycytidine" Toxics 10, no. 2: 100. https://doi.org/10.3390/toxics10020100