A Rapid Method for the Identification of Fresh and Processed Pagellus erythrinus Species against Frauds


 ,
,  ,
, 
Abstract
:
1. Introduction
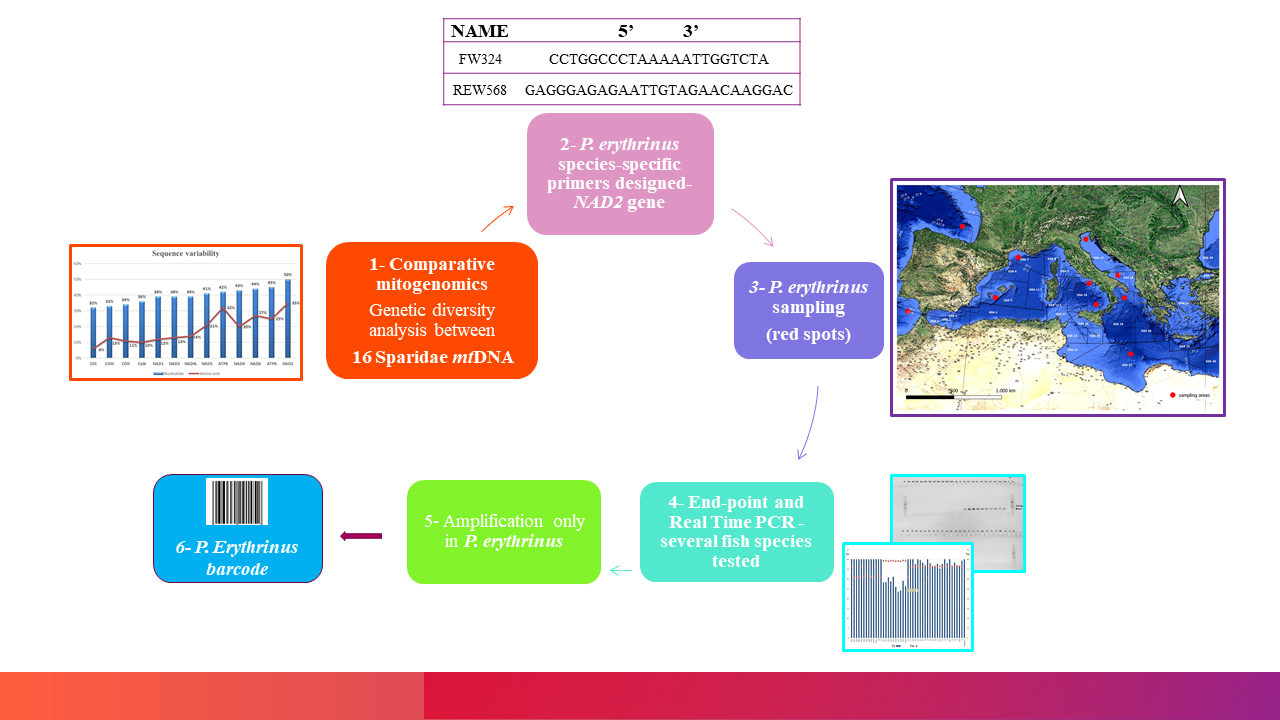
2. Materials and Methods
2.1. Fish Samples
2.2. Total Genomic DNA Extraction
2.3. Comparative Analysis of mtDNA Complete Sequences
2.4. NAD2 Fragment Amplification and Sequence Analysis
2.5. Real-Time PCR
3. Results
3.1. Pagellus erythrinus mtDNA Comparative Data
3.2. NAD2 Amplification and Analysis
3.3. NAD2 Amplification by RT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Spink, J.; Embarek, P.B.; Savelli, C.J.; Bradshaw, A. Global perspectives on food fraud: Results from a WHO survey of members of the International Food Safety Authorities Network (INFOSAN). Npj Sci. Food 2019, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Spink, J.; Moyer, D.C. Defining the public health threat of food fraud. J. Food Sci. 2011, 76, R157–R162. [Google Scholar] [CrossRef] [PubMed]
- Spink, J. The current state of food fraud prevention: Overview and requirements to address “How to Start?” and “How Much is Enough?”. Curr. Opin. Food Sci. 2019, 27, 130–138. [Google Scholar] [CrossRef]
- Lang, R.E.D. Report on the Food Crisis, Fraud in the Food Chain and the Control Thereof. Tech. Report No. 2013/2091(INI); Committee on the Environment, Public Health and Food Safety, European Parliament: Strasbourg, France, 2013. [Google Scholar]
- Chen, J.; CFSA. Chinese National Center for Food Safety Risk Assessment. Strategies for ensuring food safety of Chinese products—Fight against food fraud. In Proceedings of the Institute for Food Technologists (IFT) Annual Conference, Chicago, IL, USA, 11–14 July 2015. [Google Scholar]
- EU Food Fraud Network and the System for Administrative Assistance. Food Fraud Annual Report 2018. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/ff_ffn_annual-report_2018.pdf (accessed on 7 August 2020).
- FAO. FishStat Plus. Available online: http://www.fao:fishery/statistics/software/fishstat/en (accessed on 7 August 2020).
- Basurco, B.; Lovatelli, A.; Garcia, B. Current status of Sparidae aquaculture. In Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; Pavlidis, A., Mylonas, C.C., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2011; pp. 10–11. [Google Scholar]
- Malandra, R.; Renon, P. Le Principali Frodi dei Prodotti della Pesca; Libreria Universitaria Valdina: Milano, Italy, 1998; pp. 89–91; 105–106. [Google Scholar]
- Antonucci, F.; Costa, C.; Aguzzi, J.; Cataudella, S. Ecomorphology of morpho-functional relationships in the family of sparidae: A quantitative statistic approach. J. Morphol. 2009, 270, 843–855. [Google Scholar] [CrossRef]
- Messina, A. Identificazione di Sparidi di Interesse Commerciale Sui Mercati Internazionali Mediante DNA Barcoding. Specialist Thesis, Università di Pisa, Pisa, Italy, 2012. [Google Scholar]
- Riina, M.V.; Gilli, M.; Costa, R.; Colussi, S.; Bertuzzi, S.; Trisorio, S.; Gili, S.; Perazzini, A.Z.; Pezzolato, M.; Richelmi, G.; et al. Evaluation of DNA Markers for Fish Identification. Ital. J. Food Saf. 2012, 1, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Armani, A.; Guardone, L.; Castigliego, L.; D’Amico, P.; Messina, A.; Malandra, R.; Gianfaldoni, D.; Guidi, A. DNA and Mini-DNA Barcoding for the Identification of Porgies Species (Family Sparidae) of Commercial Interest on the International Market. Food Control 2015, 50, 589–596. [Google Scholar] [CrossRef]
- Trotta, M.; Schönhuth, S.; Pepe, T.; Cortesi, M.L.; Puyet, A.; Bautista, J.M. Multiplex PCR Method for use in real-time PCR for identification of fish fillets from grouper (Epinephelus and Mycteroperca Species) and common substitute species. J. Agric. Food Chem. 2005, 53, 2039–2045. [Google Scholar] [CrossRef]
- Pepe, T.; Trotta, M.; di Marco, I.; Anastasio, A.; Bautista, J.M.; Cortesi, M.L. Fish species identification in surimi-based products. J. Agric. Food Chem. 2007, 55, 3681–3685. [Google Scholar] [CrossRef]
- Cawthorn, D.M.; Steinman, H.A.; Witthuhn, R.C. Evaluation of the 16S and 12S RRNA genes as universal markers for the identification of commercial fish species in South Africa. Gene 2012, 491, 40–48. [Google Scholar] [CrossRef]
- Melo, B.F.; Dorini, B.F.; Foresti, F.; Oliveira, C. Little divergence among mitochondrial lineages of Prochilodus (Teleostei, Characiformes). Front. Genet. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Yang, H.; Xie, Z.; Yang, X.; Xiao, L.; Li, S.; Chen, M.; Zhao, H.; Zhang, Y. The complete mitochondrial genome of the Rhabdosargus Sarba (Perciformes: Sparidae). Mitochondrial DNA Part A 2014, 27, 1606–1607. [Google Scholar] [CrossRef]
- Chagas, A.T.D.A.; Ludwig, S.; Pimentel, J.D.S.M.; De Abreu, N.L.; Rodriguez, D.N.; Leal, H.G.; Kalapothakis, E. Use of complete mitochondrial genome sequences to identify barcoding markers for groups with low genetic distance. Mitochondrial DNA Part A 2020, 31, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Ceruso, M.; Mascolo, C.; Anastasio, A.; Pepe, T.; Sordino, P. Frauds and fish species authentication: study of the complete mitochondrial genome of some sparidae to provide specific barcode markers. Food Control 2019, 103, 36–47. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of Biochemistry: Life at the Molecular Level, 4th ed.; Wiley: Hoboken, NJ, USA, 2013; pp. 581–620. [Google Scholar]
- Abbas, B.; Tjolli, I.; Dailami, M. Phylogenetic of sago palm (Metroxylon sagu) and others monocotyledon based on mitochondrial nad2 gene markers. Biodiversitas J. Biol. Divers. 2019, 20, 2249–2256. [Google Scholar] [CrossRef] [Green Version]
- Vanhove, M.P.M.; Briscoe, A.G.; Jorissen, M.W.P.; Littlewood, D.T.J.; Huyse, T. The first next-generation sequencing approach to the mitochondrial phylogeny of African monogenean parasites (Platyhelminthes: Gyrodactylidae and Dactylogyridae). BMC Genom. 2018, 19, 520. [Google Scholar] [CrossRef]
- Feng, X.; Johnson, E.G.; Williams, E.P.; Place, A.R. Successful identification and discrimination of hatchery-reared blue crabs (Callinectes sapidus) released into the chesapeake bay using a genetic tag. J. Shellfish. Res. 2017, 36, 277–282. [Google Scholar] [CrossRef]
- Zhang, D.; Zou, H.; Wu, S.G.; Li, M.; Jakovlić, I.; Zhang, J.; Chen, R.; Li, W.X.; Wang, G.T. Three new Diplozoidae mitogenomes expose unusual compositional biases within the Monogenea class: Implications for phylogenetic studies. BMC Evol. Biol. 2018, 18, 133. [Google Scholar] [CrossRef] [Green Version]
- Ceruso, M.; Venuti, I.; Osca, D.; Caputi, L.; Anastasio, A.; Crocetta, F.; Sordino, P.; Pepe, T. The complete mitochondrial genome of the sharpsnout seabream diplodus puntazzo (Perciformes : Sparidae). Mitochondrial DNA Part B 2020, 5, 2379–2381. [Google Scholar] [CrossRef]
- Mascolo, C.; Ceruso, M.; Sordino, P.; Palma, G.; Anastasio, A.; Pepe, T. Comparison of mitochondrial DNA enrichment and sequencing methods from fish tissue. Food Chem. 2019, 294, 333–338. [Google Scholar] [CrossRef]
- Kumar, A.; Chordia, N. Methods in molecular biology. In Silico PCR Primer Designing and Validation; Basu, C., Ed.; Humana Press: New York, NY, USA, 2015; Volume 1275, pp. 143–151. [Google Scholar] [CrossRef]
- Mitchell, M. An Introduction to Genetic Algorithms; MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Xia, K.; Jiang, S. Complete mitochondrial DNA sequence of the yellowfin seabream Acanthopagrus latus and a genomic comparison among closely related sparid species. Mitochondrial DNA 2008, 19, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Su, Y.; Wang, J.; Ding, S.; Mao, Y. Complete mitochondrial genome of black porgy Acanthopagrus schlegelii (Perciformes, Sparidae). Mitochondrial DNA 2012, 23, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Mascolo, C.; Ceruso, M.; Chirollo, C.; Palma, G.; Anastasio, A.; Sordino, P.; Pepe, T. The complete mitochondrial genome of the Angolan dentex Dentex angolensis (Perciformes: Sparidae). Mitochondrial DNA Part B 2019, 4, 1245–1246. [Google Scholar] [CrossRef] [Green Version]
- Ceruso, M.; Mascolo, C.; Palma, G.; Anastasio, A.; Pepe, T.; Sordino, P. The complete mitochondrial genome of the common dentex, Dentex dentex (Perciformes: Sparidae). Mitochondrial DNA Part B 2018, 3, 391–392. [Google Scholar] [CrossRef] [Green Version]
- Mascolo, C.; Ceruso, M.; Palma, G.; Anastasio, A.; Sordino, P.; Pepe, T. The complete mitochondrial genome of the Pink dentex Dentex gibbosus (Perciformes: Sparidae). Mitochondrial DNA Part B 2018, 3, 525–526. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Jiang, L.; Xu, M.; Wu, C. Complete mitochondrial genome of yellowback sea bream Evynnis tumifrons. Mitochondrial DNA Part B 2016, 1, 352–354. [Google Scholar] [CrossRef] [Green Version]
- Mascolo, C.; Ceruso, M.; Palma, G.; Anastasio, A.; Sordino, P.; Pepe, T. The complete mitochondrial genome of the axillary seabream, Pagellus acarne (Perciformes: Sparidae). Mitochondrial DNA Part B 2018, 3, 434–435. [Google Scholar] [CrossRef]
- Ponce, M.; Infante, C.; Jimenez-Cantizano, R.M.; Perez, L.; Manchado, M. Complete mitochondrial genome of the blackspot seabream, Pagellus bogaraveo (Perciformes: Sparidae), with high levels of length heteroplasmy in the WANCY region. Gene 2008, 409, 44–52. [Google Scholar] [CrossRef]
- Ceruso, M.; Mascolo, C.; Lowe, E.K.; Palma, G.; Anastasio, A.; Pepe, T.; Sordino, P. The complete mitochondrial genome of the common Pandora Pagellus erythrinus (Perciformes: Sparidae). Mitochondrial DNA Part B 2018, 3, 624–625. [Google Scholar] [CrossRef] [Green Version]
- Ponce, M.; Infante, C.; Catanese, G.; Cardenas, S.; Manchado, M. Complete mitochondrial DNA of redbanded seabream (Pagrus auriga). In Proceedings of the Conference EAS - Aquaculture Europe 04, Barcelona, Spain, 1 September 2004. [Google Scholar]
- Afriyie, G.; Guo, Y.; Kuebutornye, F.K.A.; Larbi, C.A.; Wang, Z. Complete mitochondrial DNA sequence analysis of Bluespotted seabream, Pagrus caeruleostictus (Perciformes: Sparidae). Mitochondrial DNA Part B 2020, 5, 1913–1914. [Google Scholar] [CrossRef] [Green Version]
- Miya, M.; Kawaguchi, A.; Nishida, M. Mitogenomic exploration of higher teleostean phylogenies: A case study for moderate-scale evolutionary genomics with 38 newly determined complete mitochondrial DNA sequences. Mol. Biol. Evol. 2001, 18, 1993–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Xia, K.; Gong, J.; Jiang, S. Complete mitochondrial DNA sequence, gene organization and genetic variation of control regions in Parargyrops edita. Fish. Sci. 2007, 73, 1042–1049. [Google Scholar] [CrossRef]
- Dray, L.M.; Neuhof, M.; Diamant, A.; Huchon, D. The complete mitochondrial genome of the gilthead seabream Sparus aurata L. (Sparidae). Mitochondrial DNA 2016, 27, 781–782. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucl. Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The barcode index number (BIN) System. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Freeman, W.M.; Walker, S.J.; Vrana, K.E. Quantitative RT-PCR: Pitfalls and potential. Biotechniques 1999, 26, 112–122, 124–125. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA Barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [Green Version]
- Waugh, J. DNA barcoding in animal species: Progress, potential and pitfalls. Bioessays 2007, 29, 188–197. [Google Scholar] [CrossRef]
- Costa-Silva, G.J.; Rodriguez, M.S.; Roxo, F.F.; Foresti, F.; Oliveira, C. Using different methods to access the difficult task of delimiting species in a complex neotropical hyperdiverse group. PLoS ONE 2015, 10, e0135075. [Google Scholar] [CrossRef]
- DeSalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Zarowiecki, M.Z.; Huyse, T.; Littlewood, D.T.J. Making the most of mitochondrial genomes—Markers for phylogeny, molecular ecology and barcodes in schistosoma (Platyhelminthes: Digenea). Int. J. Parasitol. 2007, 37, 1401–1418. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, L.; Zhang, Q.; Zhou, M.; Wang, X.; Yang, X.-Z.; Yuan, M.-L. Comparative mitogenomic analysis of mirid bugs (Hemiptera: Miridae) and evaluation of potential DNA barcoding markers. PeerJ 2017, 5, e3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.N.; Iwatsuki, Y.; Yoshino, T.; Hanzawa, N. Comprehensive phylogeny of the family Sparidae (Perciformes: Teleostei) inferred from mitochondrial gene analyses. Genes Genet. Syst. 2009, 84, 153–170. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. erythrinus | |||
|---|---|---|---|
| Abbreviation | Latitude | Longitude | Sampling Areas |
| Pe1 | 38.714589 | 2.230898 | FAO 37 area—GSA5—Balearic Island |
| Pe2 | 42.774369 | 4.384218 | FAO 37 area—GSA7—Gulf of Lions |
| Pe3 | 40.775480 | 13.603978 | FAO 37 area—GSA10—South Tyrrhenian |
| Pe4 | 38.748871 | 14.524641 | FAO 37 area—GSA10—Central Tyrrhenian |
| Pe5 | 44.991156 | 13.065691 | FAO 37 area—GSA17—Northern Adriatic |
| Pe6 | 41.533674 | 17.278337 | FAO 37 area—GSA18—Southern Adriatic Sea |
| Pe7 | 39.343228 | 17.981462 | FAO 37 area—GSA19—Western Ionian Sea |
| Pe8 | 33.952939 | 18.508806 | FAO 37 area—GSA21—Southern Ionian Sea |
| Pe9 | 44.708601 | −3.470325 | FAO 27 area—Atlantic Ocean |
| Pe10 | 35.916767 | −7.425403 | FAO 34 area—Atlantic Ocean |
| Scientific Name | Family | Common Name | Abbreviation |
|---|---|---|---|
| Dentex dentex | Sparidae | Common dentex | Dd |
| Dentex gibbosus | Sparidae | Pink dentex | Dg |
| Cheimerius nufar | Sparidae | Santer seabream | Cn |
| Pagellus acarne | Sparidae | Axillary seabream | Pa |
| Aulopus filamentosus | Aulopidae | Royal flagfin | Af |
| Boops boops | Sparidae | Bogue | Bb |
| Trisopterus minutus | Gadidae | Poor cod | Tm |
| Cepola macrophthalma | Cepolidae | Red bandfish | Cm |
| Diplodus annularis | Sparidae | Annular seabream | Da |
| Diplodus sargus | Sparidae | White seabream | Ds |
| Coris julis | Labridae | Rainbow wrasse | Cj |
| Chelidonichthys lucerna | Triglidae | Tub gurnard | Cl |
| Lithognathus mormyrus | Sparidae | Sand steenbras | Lm |
| Xyrichtys novacula | Labridae | Pearly razorfish | Xn |
| Pleuronectes platessa | Pleuronectidae | European plaice | Pp |
| Lophius piscatorius | Lophiidae | Angler (= Monk) | Lp |
| Scophthalmus maximus | Scophthalmidae | Turbot | Pm |
| Sebastes capensis | Sebastidae | Cape redfish | Sc |
| Solea solea | Soleidae | Common sole | Ss |
| Trachurus trachurus | Carangidae | Atlantic horse mackerel | Tt |
| Spondyliosoma cantharus | Sparidae | Black seabream | Spc |
| Thunnus albacares | Scombridae | Yellowfin tuna | Ta |
| Thunnus obesus | Scombridae | Bigeye tuna | To |
| Thunnus thynnus | Scombridae | Atlantic bluefin tuna | Tt |
| Mullus barbatus | Mullidae | Red mullet | Mb |
| Arnoglossus laterna | Bothidae | Mediterranean scaldfish | Al |
| Dentex Dentex | |||
|---|---|---|---|
| Abbreviation | Latitude | Longitude | Sampling Areas |
| Dd1 | 39.808662 | 3.740905 | GSA 5—Balearic Island |
| Dd2 | 42.519035 | 3.534141 | GSA 7—Gulf of Lions |
| Dd3 | 38.771348 | 14.946960 | GSA 10—South Tyrrhenian |
| Dd4 | 40.409446 | 13.798889 | GSA 10—Central Tyrrhenian |
| Dd5 | 40.044575 | 9.841622 | GSA 11.2—Sardinia (East) |
| Dd6 | 44.980473 | 13.137520 | GSA 17—Northern Adriatic |
| Dd7 | 40.668284 | 18.290108 | GSA 18—Southern Adriatic Sea |
| Dd8 | 40.044587 | 17.147530 | GSA 19—Western Ionian Sea |
| Dd9 | 31.475938 | 18.685467 | GSA 21—Southern Ionian Sea |
| Dd10 | 35.338496 | 35.708826 | GSA 27—Levante |
| Dd11 | 40.874620 | 12.985808 | GSA 10—Ponza Island, Central Thyrrhenian |
| Species | Ac. Number | FAO Fishing Areas | References |
|---|---|---|---|
| Acanthopagrus latus | NC_010977 | FAO 71 | [32] |
| Acanthopagrus schlegelii | JQ_746035 | FAO 71 | [33] |
| Dentex angolensis | NC_044097 | FAO 47 | [34] |
| Dentex dentex | MG_727892 | FAO 37 | [35] |
| Dentex gibbosus | MG_653593 | FAO 37 | [36] |
| Dentex tumifrons | NC_029479 | FAO 71 | [37] |
| Diplodus puntazzo | MT319027 | FAO 37 | [26] |
| Pagellus acarne | MG_736083 | FAO 37 | [38] |
| Pagellus bogaraveo | NC_009502 | FAO 27 | [39] |
| Pagellus erythrinus | MG_653592 | FAO 37 | [40] |
| Pagrus auriga | AB_124801 | FAO 37 | [41] |
| Pagrus caeruleostictus | MN319701 | FAO 34 | [42] |
| Pagrus major | NC_003196 | Farmed and supplied from Andalusia (Spain) fish market | [43] |
| Parargyrops edita | EF_107158 | FAO 71 | [44] |
| Rhabdosargus sarba | KM_272585 | Farmed Daya Bay Aquaculture Center, Guangdong (China) | [18] |
| Sparus aurata | LK_022698 | Farmed and supplied from Jaffa (Israel) fish market | [45] |
| Primer Name | 5′→3′ Sequence | Tm °C | CG% | nt | A | T | C | G |
|---|---|---|---|---|---|---|---|---|
| FW303 | CCTGGCCCTAAAAATTGGTCTA | 65.1 | 45.5 | 22 | 6 | 6 | 6 | 4 |
| REV593 | GAGGGAGAGAATTGTAGAACAAGGAC | 65.4 | 46.2 | 26 | 11 | 3 | 2 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceruso, M.; Mascolo, C.; De Luca, P.; Venuti, I.; Smaldone, G.; Biffali, E.; Anastasio, A.; Pepe, T.; Sordino, P. A Rapid Method for the Identification of Fresh and Processed Pagellus erythrinus Species against Frauds. Foods 2020, 9, 1397. https://doi.org/10.3390/foods9101397
Ceruso M, Mascolo C, De Luca P, Venuti I, Smaldone G, Biffali E, Anastasio A, Pepe T, Sordino P. A Rapid Method for the Identification of Fresh and Processed Pagellus erythrinus Species against Frauds. Foods. 2020; 9(10):1397. https://doi.org/10.3390/foods9101397
Chicago/Turabian StyleCeruso, Marina, Celestina Mascolo, Pasquale De Luca, Iolanda Venuti, Giorgio Smaldone, Elio Biffali, Aniello Anastasio, Tiziana Pepe, and Paolo Sordino. 2020. "A Rapid Method for the Identification of Fresh and Processed Pagellus erythrinus Species against Frauds" Foods 9, no. 10: 1397. https://doi.org/10.3390/foods9101397