
UHPLC-OrbiTrap MS Characterization of Phenolic Profiles in French Marigold Extracts and Analysis of Their Antifeedant Activity against Colorado Potato Beetle

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
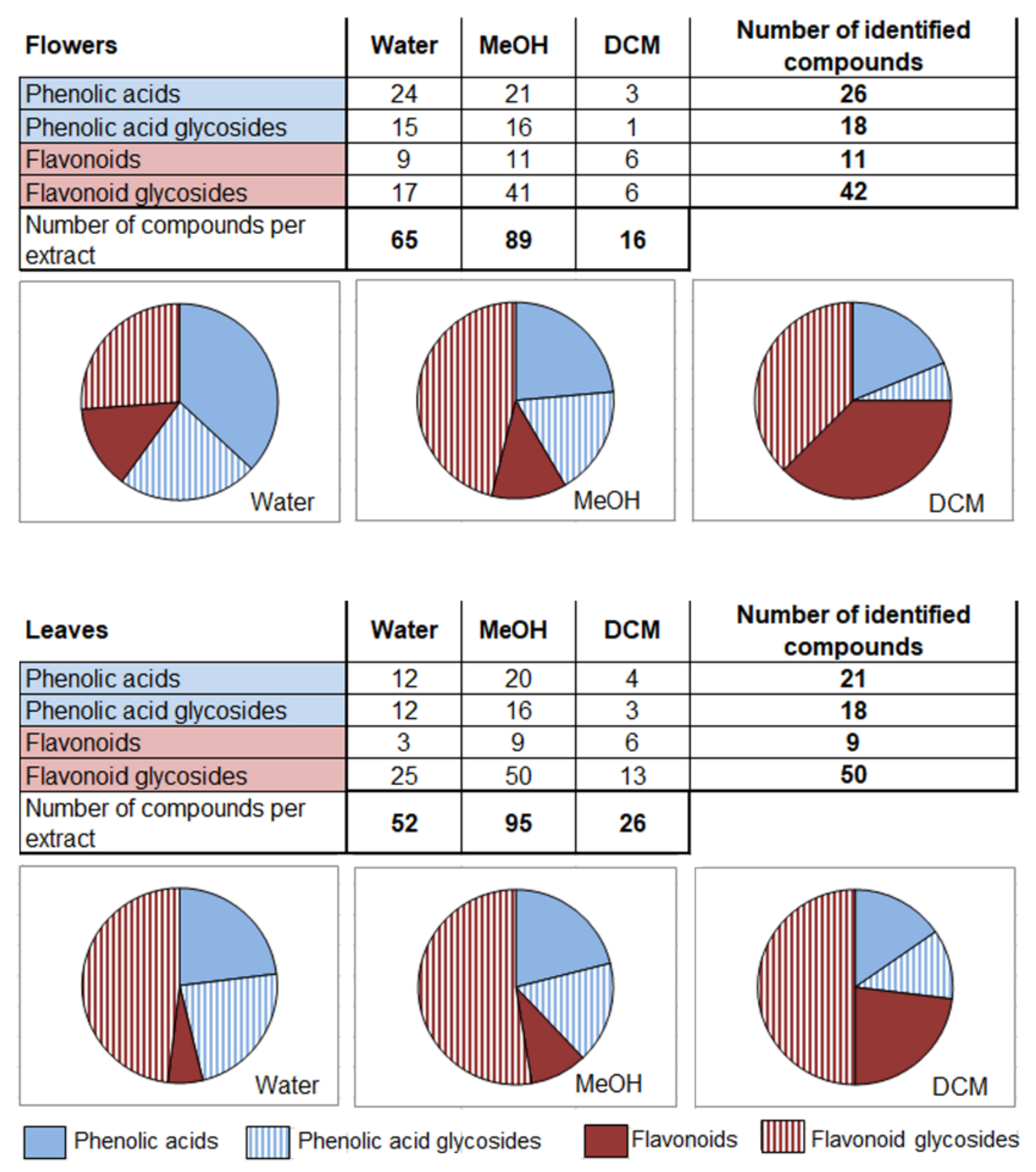
2.1. Phytochemical Profile of French Marigold Extracts
2.2. Antioxidant Activity of French Marigold Extracts
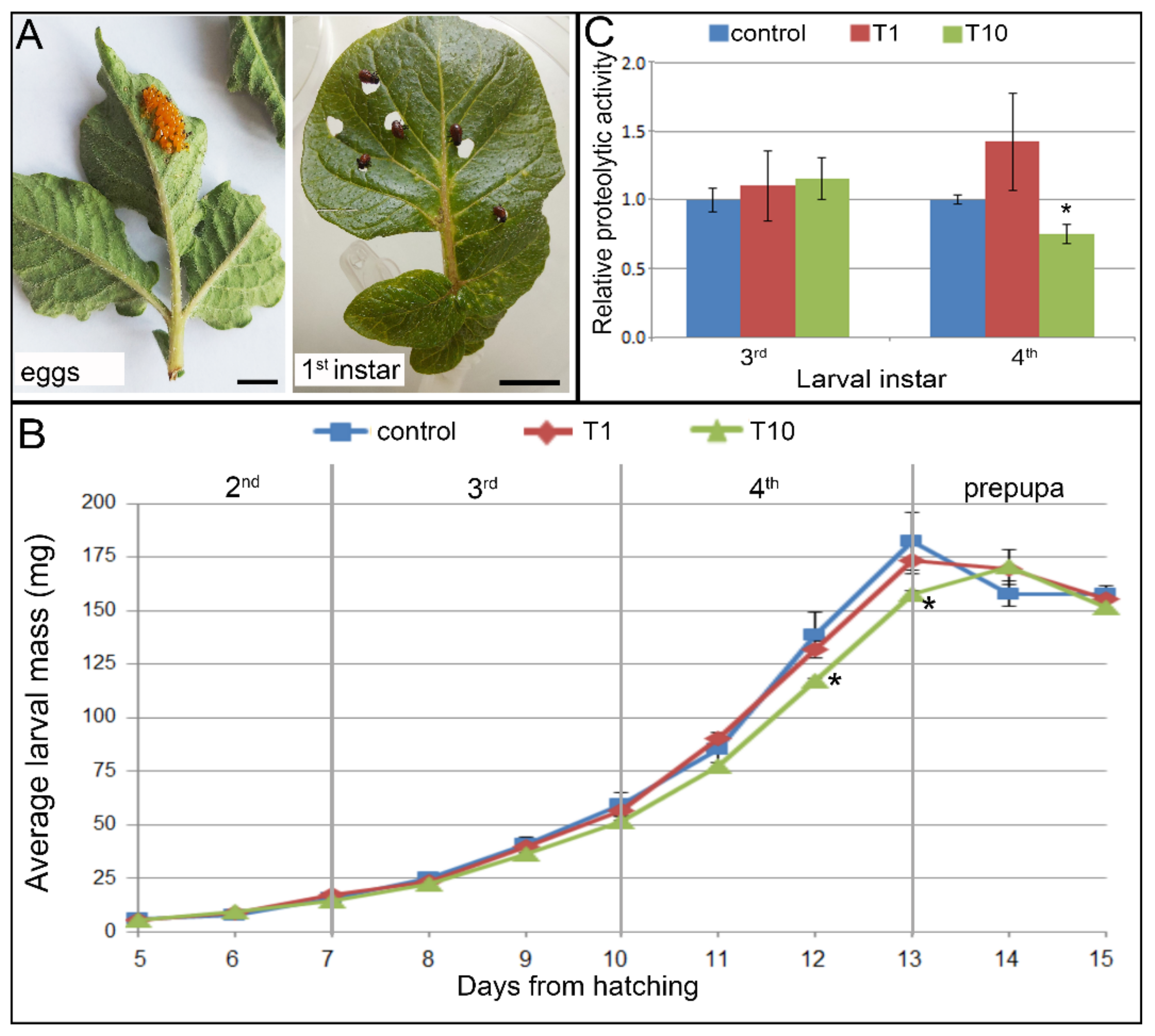
2.3. Effect of French Marigold Extract on Colorado Potato Beetle Larvae
2.3.1. Growth and Development of CPB Larvae
2.3.2. Digestive Proteolysis in Larvae Fed with French Marigold Extract-Coated Potato Leaves
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extracts Preparation
4.2. UHPLC-OrbiTrap MS Assessment of French Marigold Extracts
4.3. Determination of Total Phenolic Content (TPC)
4.4. Estimation of In Vitro Antioxidant Potential of French Marigold Extracts
4.4.1. DPPH Assay
4.4.2. ABTS Assay
4.4.3. FRAP Assay
4.5. Antifeedant Bioassay with Colorado Potato Beetle
4.5.1. Insect Rearing and Feeding with Potato Leaves Coated with French Marigold Extract
4.5.2. CPB Growth and Development
4.5.3. CPB Protease Activity Assay
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2020. Transforming Food Systems for Affordable Healthy Diets; FAO: Rome, Italy, 2020. [Google Scholar]
- FAOSTAT. Statistical Databases. Food and Agricultural Organization of the United Nations, Rome. Available online: https://www.fao.org/faostat/en/#data/RP (accessed on 21 May 2021).
- Jennings, A.A.; Li, Z. Scope of the worldwide effort to regulate pesticide contamination in surface soils. J. Environ. Manag. 2014, 146, 420–443. [Google Scholar] [CrossRef] [PubMed]
- Lai, W. Pesticide use and health outcomes: Evidence from agricultural water pollution in China. J. Environ. Econ. Manag. 2017, 86, 93–120. [Google Scholar] [CrossRef]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid pesticide residues in the global environment: An overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Costello, S.; Cockburn, M.; Zhang, X.; Bronstein, J.; Ritz, B. Parkinson’s disease risk from ambient exposure to pesticides. Eur. J. Epidemiol. 2011, 26, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Boedeker, W.; Watts, M.; Clausing, P.; Marquez, E. The global distribution of acute unintentional pesticide poisoning: Estimations based on a systematic review. BMC Public Health 2020, 20, 1875. [Google Scholar] [CrossRef]
- Cingel, A.; Savić, J.; Lazarević, J.; Ćosić, T.; Raspor, M.; Smigocki, A.; Ninković, S. Extraordinary adaptive plasticity of Colorado potato beetle: “Ten-striped Spearman” in the era of biotechnological warfare. Int. J. Mol. Sci. 2016, 17, 1538. [Google Scholar] [CrossRef]
- Gupta, S.; Dikshit, A.K. Biopesticides: An ecofriendly approach for pest control. J. Biopestic. 2010, 3, 186. [Google Scholar]
- AlSalhi, M.S.; Elumalai, K.; Devanesan, S.; Govindarajan, M.; Krishnappa, K.; Maggi, F. The aromatic ginger Kaempferia galanga L. (Zingiberaceae) essential oil and its main compounds are effective larvicidal agents against Aedes vittatus and Anopheles maculatus without toxicity on the non-target aquatic fauna. Ind. Crops Prod. 2020, 158, 113012. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Kiokias, S.; Proestos, C.; Oreopoulou, V. Phenolic acids of plant origin—A review on their antioxidant activity in vitro (o/w emulsion systems) along with their in vivo health biochemical properties. Food 2020, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Okwute, S.K. Plants as potential sources of pesticidal agents: A review. In Pesticides–Advances in Chemical and Botanical Pesticides; Soundararajan, R.P., Ed.; InTech Open: London, UK, 2012; Volume 10, pp. 208–232. [Google Scholar]
- Abdelkhalek, A.; Salem, M.Z.; Kordy, A.M.; Salem, A.Z.; Behiry, S.I. Antiviral, antifungal, and insecticidal activities of Eucalyptus bark extract: HPLC analysis of polyphenolic compounds. Microb. Pathog. 2020, 147, 104383. [Google Scholar] [CrossRef] [PubMed]
- Koul, O. Phytochemicals and insect control: An antifeedant approach. Crit. Rev. Plant Sci. 2008, 27, 1–24. [Google Scholar] [CrossRef]
- Pavela, R. The feeding effect of polyphenolic compounds on the Colorado potato beetle (Leptinotarsa decemlineata Say). Pest Technol. 2007, 1, 81–84. [Google Scholar]
- Pavela, R. Larvicidal effects of some Euro-Asiatic plants against Culex quinquefasciatus Say larvae (Diptera: Culicidae). Parasitol. Res. 2009, 105, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Rani, A.S.; Murty, U.S. Antifeedant activity of Spilanthes acmella flower head extract against Spodoptera litura (Fabricius). J. Entomol. Res. 2009, 33, 55–57. [Google Scholar]
- Mares, D.; Tosi, B.; Romagnoli, C.; Poli, F. Antifungal activity of Tagetes patula extracts. Pharm. Biol. 2002, 40, 400–404. [Google Scholar] [CrossRef]
- Dharmagadda, V.S.S.; Naik, S.N.; Mittal, P.K.; Vasudevan, P. Larvicidal activity of Tagetes patula essential oil against three mosquito species. Bioresour. Technol. 2005, 96, 1235–1240. [Google Scholar] [CrossRef]
- Rondon, M.; Velasco, J.; Hernandez, J.; Pecheneda, M.; Rojas, J.; Morales, A.; Carmona, J.; Diaz, T. Chemical composition and antibacterial activity of the essential oil of Tagetes patula L. (Asteraceae) collected from the Venezuela Andes. Rev. Latinoam. Quím. 2006, 34, 32. [Google Scholar]
- Politi, F.A.S.; Nascimento, J.D.; da Silva, A.A.; Moro, I.J.; Garcia, M.L.; Guido, R.V.C.; Pietro, R.C.L.R.; Godinho, A.F.; Furlan, M. Insecticidal activity of an essential oil of Tagetes patula L. (Asteraceae) on common bed bug Cimex lectularius L. and molecular docking of major compounds at the catalytic site of ClAChE1. Parasitol. Res. 2017, 116, 415–424. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Yool, A.J.; Spurgeon, D.W. Insecticidal activity of marigold Tagetes patula plants and foliar extracts against the hemipteran pests, Lygus hesperus and Bemisia tabaci. PLoS ONE 2020, 15, e0233511. [Google Scholar] [CrossRef] [PubMed]
- Politi, F.; Queiroz-Fernandes, G.M.; Rodrigues, E.R.; Freitas, J.A.; Pietro, R. Antifungal, antiradical and cytotoxic activities of extractives obtained from Tagetes patula L. (Asteraceae), a potential acaricide plant species. Microb. Pathog. 2016, 95, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, D.K.; Bisht, M.S.; Uain, S.C.; Metha, C.K.; Sharma, G.C. Quercetagetin 5-methyl ether from the petals of Tagetes patula. Phytochemistry 1980, 19, 713–714. [Google Scholar] [CrossRef]
- Ivancheva, S.; Zdravkova, M. Flavonoids in Tagetes patula. Fitoterapia 1993, 64, 555–558. [Google Scholar]
- Vasudevan, P.; Kashyap, S.; Sharma, S. Tagetes: A multipurpose plant. Bioresour. Technol. 1997, 62, 29–35. [Google Scholar] [CrossRef]
- Faizi, S.; Dar, A.; Siddiqi, H.; Naqvi, S.; Naz, A.; Bano, S.; Lubna. Bioassay-guided isolation of antioxidant agents with analgesic properties from flowers of Tagetes patula. Pharm. Biol. 2011, 49, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Faizi, S.; Fayyaz, S.; Bano, S.; Iqbal, E.Y.; Lubna; Siddiqi, H.; Naz, A. Isolation of nematicidal compounds from Tagetes patula L. yellow flowers: Structure-activity relationship studies against cyst nematode Heterodera zeae infective stage larvae. J. Agr. Food Chem. 2011, 59, 9080–9093. [Google Scholar] [CrossRef]
- Munhoz, M.V.; Longhini, R.; Souza, R.P.J.; Zequi, A.C.J.; Leite Mello, V.S.E.; Lopes, G.C.; Mello, C.P.J. Extraction of flavonoids from Tagetes patula: Process optimization and screening for biological activity. Rev. Bras. Farmacogn. 2014, 24, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Perich, M.J.; Wells, C.; Bertsch, W.; Tredway, K.E. Toxicity of extracts from three Tagetes against adults and larvae of yellowfever mosquito and Anopheles stephensi (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 833–837. [Google Scholar] [CrossRef]
- Leger, C.; Riga, E. Evaluation of marigolds and entomopathogenic nematodes for control of the cabbage maggot Delia radicum. J. Sustain. Agric. 2009, 33, 128–141. [Google Scholar] [CrossRef]
- Forister, M.L.; Ehmer, A.G.; Futuyma, D.J. The genetic architecture of a niche: Variation and covariation in host use traits in the Colorado potato beetle. J. Evol. Biol. 2007, 20, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Alyokhin, A.; Vincent, C.; Giordanengo, P. Insect Pests of Potato: Global Perspectives on Biology and Management, 1st ed.; Academic Press: Oxford, UK, 2013; p. 613. [Google Scholar]
- Arthropod Pesticide Resistance Database (APRD). Leptinotarsa decemlineata—Shown Resistance to Active Ingredient(s). Available online: https://www.pesticideresistance.org/display.php?page=species&arId=141 (accessed on 1 September 2021).
- Gauthier, N.L.; Hofmaster, R.N.; Semel, M. History of Colorado potato beetle control. Adv. Potato Pest Manag. 1981, 23, 13–33. [Google Scholar]
- Bina, S.; Javadi, I.; Iravani, O. Evaluation of the repellency effect of neem (Melia azedarach) plant extracts based on the Mittler & Dadd method. J. Agric. Chem. Environ. 2017, 6, 165–174. [Google Scholar]
- Moreau, T.L.; Warman, P.R.; Hoyle, J. An evaluation of companion planting and botanical extracts as alternative pest controls for the Colorado potato beetle. Biol. Agric. Hortic. 2006, 23, 351–370. [Google Scholar] [CrossRef]
- Mohammed, F.; Gorla, M.; Bisoyi, V.; Tammineni, P.; Babu, N.; Sepuri, V.N. Rotenone-induced reactive oxygen species signal the recruitment of STAT3 to mitochondria. FEBS Lett. 2020, 594, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Tanner, C.M.; Kamel, F.; Webster, R.G.; Hoppin, J.A.; Goldman, S.M.; Korell, M.; Marras, C.; Bhudhikanok, G.S.; Kasten, M.; Chade, A.R.; et al. Rotenone, Paraquat and Parkinson’s disease. Environ. Health Perspect. 2011, 119, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Gökçe, A.; Whalon, M.E.; Çam, H.; Yanar, Y.; Demirtaş, I.; Gőren, N. Contact and residual toxicities of 30 plant extracts to Colorado potato beetle larvae. Arch. Phytopathol. Pflanzenschutz. 2007, 6, 441–450. [Google Scholar] [CrossRef]
- Ferreres, F.; Llorach, R.; Gil-Izquierdo, A. Characterization of the interglycosidic linkage in di-, tri-, tetra- and pentaglycosylated flavonoids and differentiation of positional isomers by liquid chromatography/electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2004, 39, 312–321. [Google Scholar] [CrossRef]
- Cuyckens, F.; Rozenberg, R.; de Hoffmann, E.; Claeys, M. Structure characterization of flavonoid O-diglycosides by positive and negative nano-electrospray ionization ion trap mass spectrometry. J. Mass Spectrom. 2001, 36, 1203–1210. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, Z.; Fang, J.; Liu, M.; Niu, Y.; Chen, S.; Wang, H. An on-line high-performance liquid chromatography-diode-array detector-electrospray ionization-ion-trap-time-of-flight-mass spectrometry-total antioxidant capacity detection system applying two antioxidant methods for activity evaluation of the edible flowers from Prunus mume. J. Chromatogr. A. 2015, 1414, 88–102. [Google Scholar]
- Ablajan, K.; Abliz, Z.; Shang, X.-Y.; He, J.-M.; Zhang, R.-P.; Shi, J.-G. Structural characterization of flavonol 3,7-di-O-glycosides and determination of the glycosylation position by using negative ion electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2006, 41, 352–360. [Google Scholar] [CrossRef] [PubMed]
- March, R.E.; Lewars, E.G.; Stadey, C.J.; Miao, X.-S.; Zhao, X.; Metcalfe, C.D. A comparison of flavonoid glycosides by electrospray tandem mass spectrometry. Int. J. Mass Spectrom. 2006, 248, 61–85. [Google Scholar] [CrossRef]
- Barchan, A.; Bakkali, M.; Arakrak, A.; Pagán, R.; Laglaoui, A. The effects of solvents polarity on the phenolic contents and antioxidant activity of three Mentha species extracts. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 399–412. [Google Scholar]
- Azwanida, N.N. A review on the extraction methods use in medicinal plants, principle, strength, and limitation. Med. Aromat. Plants 2015, 4, 196. [Google Scholar]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Bhave, A.; Schulzová, V.; Mrnka, L.; Hajšlová, J. Influence of harvest date and postharvest treatment on carotenoid and flavonoid composition in French marigold flowers. J. Agric. Food Chem. 2020, 68, 7880–7889. [Google Scholar] [CrossRef]
- Politi, F.A.S.; Figueira, G.M.; Araújo, A.M.; Sampieri, B.R.; Mathias, M.I.; Szabó, M.P.; Bechara, G.H.; Dos Santos, L.C.; Vilegas, W.; Pietro, R.C. Acaricidal activity of ethanolic extract from aerial parts of Tagetes patula L. (Asteraceae) against larvae and engorged adult females of Rhipicephalus sanguineus (Latreille, 1806). Parasites Vectors 2012, 5, 295. [Google Scholar] [CrossRef] [Green Version]
- Hvattum, E.; Ekeberg, D. Study of the collision-induced radical cleavage of flavonoid glycosides using negative electrospray ionization tandem quadrupole mass spectrometry. J. Mass Spectrom. 2003, 38, 43–49. [Google Scholar] [CrossRef]
- Hammoda, H.M. Flavonoids from the flowers of Tagetes erecta L. Alex. J. Pharm. Sci. 2004, 18, 93–96. [Google Scholar]
- Mohamed, G.A. Tagenols A and B: New lipoxygenase inhibitor flavonols from Tagetes minuta. Phytochem. Lett. 2016, 16, 141–145. [Google Scholar] [CrossRef]
- Farkas, O.; Jakus, J.; Héberger, K. Quantitative structure—Antioxidant activity relationships of flavonoid compounds. Molecules 2004, 9, 1079–1088. [Google Scholar]
- Rodriguez-Bonilla, P.; Gandia-Herrero, F.; Matencio, A.; Garcia-Carmona, F.; Lopez-Nicolas, J. Comparative study of the antioxidant capacity of four stilbenes using ORAC, ABTS+, and FRAP techniques. Food Anal. Method. 2017, 10, 2994–3000. [Google Scholar] [CrossRef]
- Farhoosh, R.; Johnny, S.; Asnaashari, M.; Molaahmadibahraseman, N.; Sharif, A. Structure—antioxidant activity relationships of o-hydroxyl, o-methoxy, and alkyl ester derivatives of p-hydroxybenzoic acid. Food Chem. 2016, 194, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shazhad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef] [PubMed]
- Kashif, M.; Bano, S.; Naqvi, S.; Faizi, S.; Lubna; Mesaik, A.; Azeemi, K.; Farooq, A. Cytotoxic and antioxidant properties of phenolic compounds from Tagetes patula flower. Pharm. Biol. 2014, 53, 1–10. [Google Scholar]
- Alvarado-Sansininea, J.; Luis Sánchez-Sánchez, L.; López-Muñoz, H.; Escobar, M.L.; Flores-Guzmán, F.; Tavera-Hernández, R.; Jiménez-Estrada, M. Quercetagetin and patuletin: Antiproliferative, necrotic and apoptotic activity in tumor cell lines. Molecules 2018, 23, 2579. [Google Scholar] [CrossRef] [Green Version]
- Faizi, S.; Siddiqi, H.; Bano, S.; Naz, A.; Lubna; Mazhar, K.; Khan, S.A. Antibacterial and antifungal activities of different parts of Tagetes patula: Preparation of patuletin derivatives. Pharm. Biol. 2008, 46, 309–320. [Google Scholar] [CrossRef]
- Jabeen, A.; Mesaik, M.A.; Simjee, S.U.; Lubna; Bano, S.; Faizi, S. Anti-TNF-α and anti-arthritic effect of patuletin: A rare flavonoid from Tagetes patula. Int. Immunopharmacol. 2016, 36, 232–240. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Barbehenn, R.; Cheek, S.; Gasperut, A.; Lister, E.; Maben, R. Phenolic compounds in red oak and sugar maple leaves have prooxidant activities in the midgut fluids of Malacosoma disstria and Orgyia leucostigma caterpillars. J. Chem. Ecol. 2005, 31, 969–988. [Google Scholar] [CrossRef] [Green Version]
- Barbehenn, R.; Dodick, T.; Poopat, U.; Spencer, B. Fenton-type reactions and iron concentrations in the midgut fluids of tree-feeding caterpillars. Arch. Insect Biochem. Physiol. 2005, 60, 32–43. [Google Scholar] [CrossRef]
- Villanueva, C.; Kross, R.D. Antioxidant-induced stress. Int. J. Mol. Sci. 2012, 13, 2091–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, S.C.; Gray, G.C. Kaempferol-induced nuclear DNA damage and lipid peroxidation. Cancer Lett. 1994, 85, 159–164. [Google Scholar] [CrossRef]
- Sahu, S.C.; Washington, M.C. Quercetin-induced lipid peroxidation and DNA damage in isolated rat-liver nuclei. Cancer Lett. 1991, 58, 75–79. [Google Scholar] [CrossRef]
- Ferro, D.N.; Logan, J.A.; Vos, R.H.; Elkinston, J.S. Colorado potato beetle temperature depended growth and feeding rates. Environ. Entomol. 1985, 14, 343–348. [Google Scholar] [CrossRef]
- Pavela, R. Antifeedant activity of plant extracts on Leptinotarsa decemlineata Say. and Spodoptera littoralis Bois. larvae. Ind. Crops Prod. 2010, 32, 213–219. [Google Scholar] [CrossRef]
- Zehnder, G.; Warthen, J.D. Feeding inhibition and mortality effects of neem-seed extract on the Colorado potato beetle (Coleoptera, Chrysomelidae). J. Econ. Entomol. 1988, 81, 1040–1044. [Google Scholar] [CrossRef]
- Igrc Barčić, J.; Bažok, R.; Bezjak, S.; Gotlin Čuljak, T.; Barčić, J. Combinations of several insecticides used for integrated control of Colorado potato beetle (Leptinotarsa decemlineata, Say, Coleoptera: Chrysomelidae). J. Pest. Sci. 2006, 79, 223. [Google Scholar] [CrossRef]
- Song, J.; Kwon, O.; Chen, S.; Daruwala, R.; Eck, P.; Park, J.B.; Levine, M. Flavonoid inhibition of sodium-dependent vitamin C transporter 1 (SVCT1) and glucose transporter isoform 2 (GLUT2), intestinal transporters for vitamin C and Glucose. J. Biol. Chem. 2002, 277, 15252–15260. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Suzuki, M.; Satsu, H.; Arai, S.; Hara, Y.; Suzuki, K.; Miyamoto, Y.; Shimizu, M. Green tea polyphenols inhibit the sodium-dependent glucose transporter of intestinal epithelial cells by a competitive mechanism. J. Agric. Food Chem. 2000, 48, 5618–5623. [Google Scholar] [CrossRef]
- Nijhout, H.F. The control of body size in insects. Dev. Biol. 2003, 261, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Devi, U.; Bora, D. Growth inhibitory effect of phenolic extracts of Ziziphus jujuba Mill. in dengue vector Aedes aegypti (L) in parent and F1 generation. Asian Pac. J. Trop. Med. 2017, 10, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kwon, C.S.; Son, K.H. Inhibition of ά-glucosidase and ά-amylase by luteolin, a flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Lokumcu Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef]
- Šuković, D.; Knežević, B.; Gašić, U.; Sredojević, M.; Ćirić, I.; Todić, S.; Mutić, J.; Tešić, Ž. Phenolic profiles of leaves, grapes and wine of grapevine variety Vranac (Vitis vinifera L.) from Montenegro. Foods 2020, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- Siriamornpun, S.; Kaisoon, O.; Meeso, N. Changes in colour, antioxidant activities and carotenoids (lycopene, β-carotene, lutein) of marigold flower (Tagetes erecta L.) resulting from different drying processes. J. Func. Food. 2012, 4, 757–766. [Google Scholar] [CrossRef]
- Wang, Y.M.; Ran, X.K.; Riaz, M.; Yu, M.; Cai, Q.; Dou, D.Q.; Metwaly, A.M.; Kang, T.G.; Cai, D.C. Chemical constituents of stems and leaves of Tagetes patula L. and its fingerprint. Molecules 2019, 24, 3911. [Google Scholar] [CrossRef] [Green Version]
- Burlec, A.F.; Pecio, Ł.; Kozachok, S.; Mircea, C.; Corciovă, A.; Vereștiuc, L.; Cioancă, O.; Oleszek, W.; Hăncianu, M. Phytochemical profile, antioxidant activity, and cytotoxicity assessment of Tagetes erecta L. flowers. Molecules 2021, 26, 1201. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use a free radical method to evaluate antioxidative activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorisation assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overney, S.; Fawe, A.; Yelle, S.; Michaud, D. Dietrelated plasticity of the digestive proteolytic system in larvae of the Colorado potato beetle (Leptinotarsa decemlineata Say). Arch. Insect Biochem. Physiol. 1997, 36, 241–250. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein using the principle of dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Michaud, D.; Nguyen-Quoc, B.; Yelle, S. Production of Oryzacystatins I and II in Escherichia coli using the glutathione S-transferase gene fusion system. Biotechnol. Prog. 1994, 10, 155–159. [Google Scholar] [CrossRef]
- Zengin, G.; Cvetanović, A.; Gašić, U.; Dragićević, M.; Stupar, A.; Uysal, A.; Şenkardes, I.; Sinan, K.I.; Picot-Allain, M.C.N.; Güneş, A.K.; et al. UHPLC-LTQ OrbiTrap MS analysis and biological properties of Origanum vulgare subsp. viridulum obtained by different extraction methods. Ind. Crops Prod. 2020, 154, 112747. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPC (mg GAE/g) | DPPH IC50 (mg/mL) | ABTS IC50 (mg/mL) | FRAP IC50 (mg/mL) | ||
|---|---|---|---|---|---|
| Flowers | Water | 37.31 ab | 0.404 a | 1.023 b | 0.120 bc |
| Methanol | 81.56 c | 0.690 a | 0.489 a | 0.044 ab | |
| Dichloromethane | 37.34 ab | 1.722 b | 2.158 c | 0.231 d | |
| Leaves | Water | 39.88 ab | 3.176 c | 1.694 bc | 0.180 c |
| Methanol | 53.00 b | 0.152 a | 1.436 bc | 0.096 ab | |
| Dichloromethane | 29.84 a | 5.465 d | 2.578 d | 0.299 d | |
| Trolox | / | 0.013 a | 0.103 a | 0.010 a | |
| R2 | / | −0.551 | −0.799 | −0.820 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devrnja, N.; Gašić, U.; Šajkunić, S.; Cingel, A.; Stupar, S.; Tubić, L.; Savić, J. UHPLC-OrbiTrap MS Characterization of Phenolic Profiles in French Marigold Extracts and Analysis of Their Antifeedant Activity against Colorado Potato Beetle. Plants 2022, 11, 407. https://doi.org/10.3390/plants11030407
Devrnja N, Gašić U, Šajkunić S, Cingel A, Stupar S, Tubić L, Savić J. UHPLC-OrbiTrap MS Characterization of Phenolic Profiles in French Marigold Extracts and Analysis of Their Antifeedant Activity against Colorado Potato Beetle. Plants. 2022; 11(3):407. https://doi.org/10.3390/plants11030407
Chicago/Turabian StyleDevrnja, Nina, Uroš Gašić, Sanja Šajkunić, Aleksandar Cingel, Sofija Stupar, Ljiljana Tubić, and Jelena Savić. 2022. "UHPLC-OrbiTrap MS Characterization of Phenolic Profiles in French Marigold Extracts and Analysis of Their Antifeedant Activity against Colorado Potato Beetle" Plants 11, no. 3: 407. https://doi.org/10.3390/plants11030407