Following the Pathway of W Chromosome Differentiation in Triportheus (Teleostei: Characiformes)


 ,
,  , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Samples and Chromosomal Preparations
2.2. Probe Labeling and Fluorescence In Situ Hybridization (FISH)
2.3. Microscopy Analysis and Image Processing
2.4. Polymerase Chain Reaction (PCR) Using the DOP-PCR Amplified Microdissected Z Chromosomes
3. Results
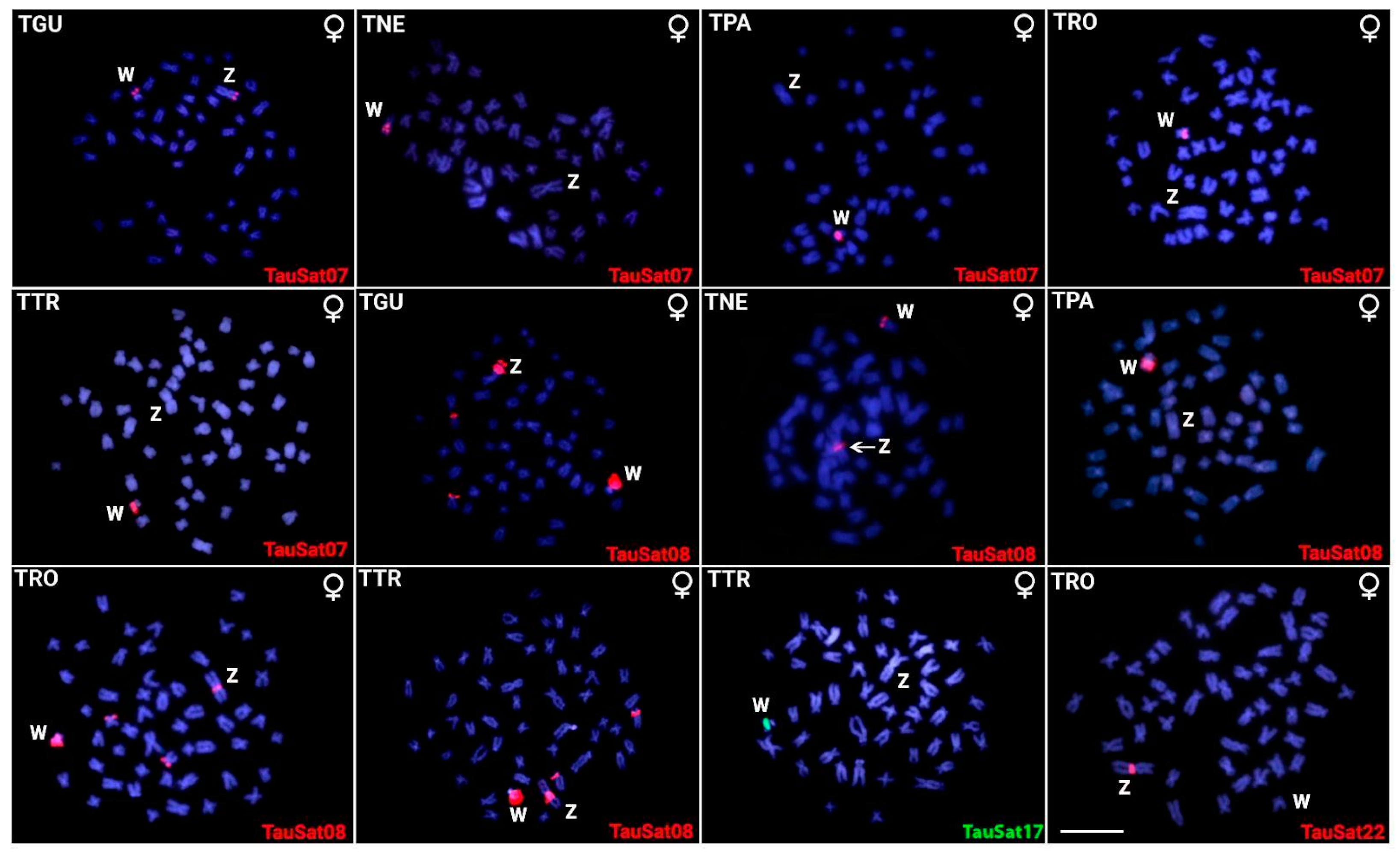
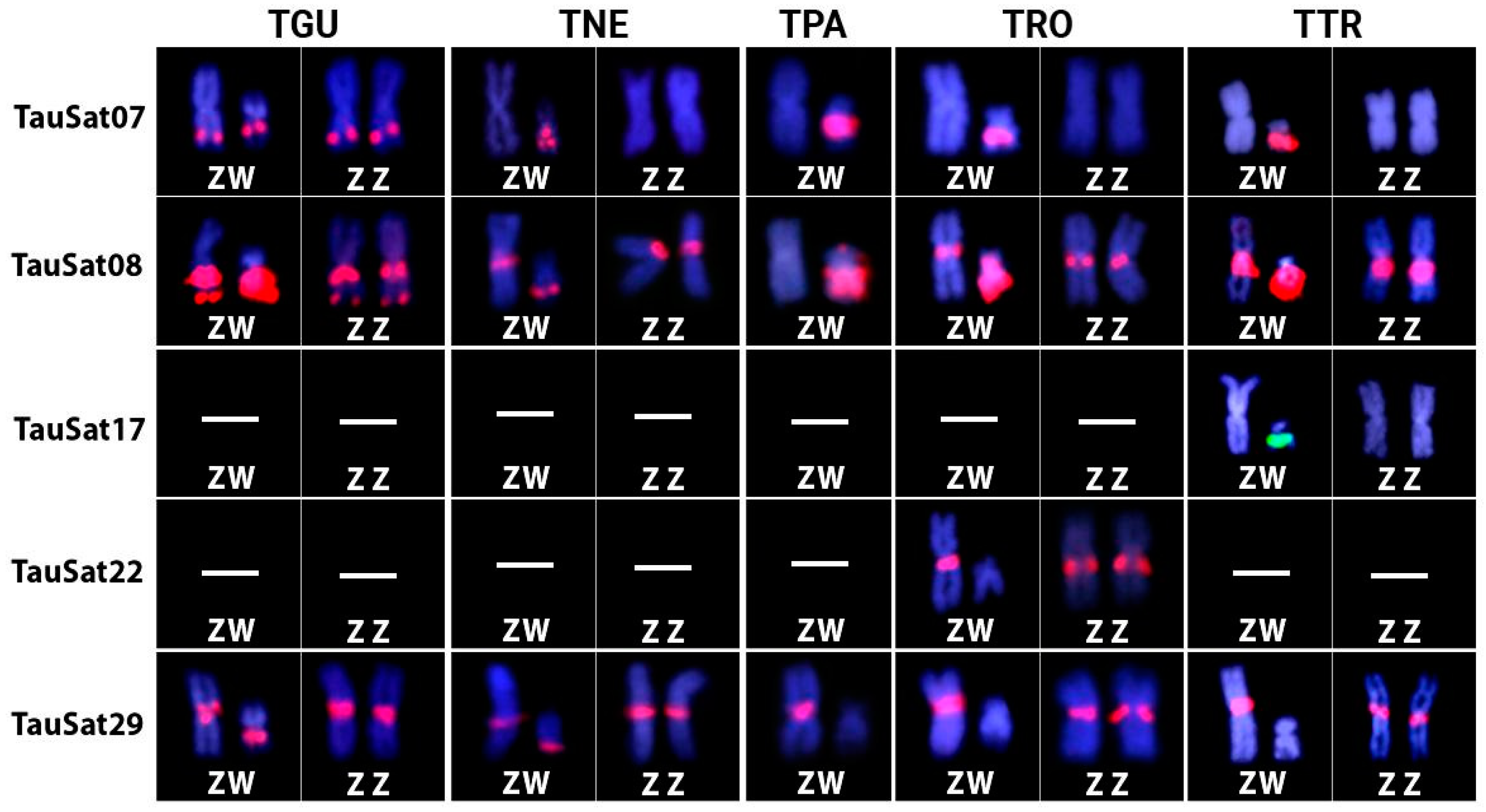
3.1. Chromosomal Location of TauSatDNAs
3.2. SatDNA Content of TAU Z Chromosome
4. Discussion
4.1. The W Chromosome Differentiation
4.2. From W to Z Chromosomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer: Berlin/Heidelberg, Germany, 1967; pp. 123–135. ISBN 978-364-288-178-7. [Google Scholar]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J. Evolution of Sex Determining Mechanisms; Benjamin/Cummings: San Francisco, CA, USA, 1983; ISBN 978-080-530-400-8. [Google Scholar]
- Bachtrog, D. A dynamic view of sex chromosome evolution. Curr. Opin. Genet. Dev. 2006, 16, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Evolution of the Y sex chromosome in animals. Bioscience 1996, 46, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Stöck, M.; Rovatsos, M.; Bullejos, M.; Herpin, A.; Jeffries, D.L.; Pokorná, M.J. Expanding the classical paradigm: What we have learnt from vertebrates about sex chromosome evolution. Philos. Trans. R. Soc. B 2021, 376, 20200097. [Google Scholar] [CrossRef]
- Graves, J.A.M.; Shetty, S. Sex from W to Z: Evolution of vertebrate sex chromosomes and sex-determining genes. J. Exp. Zool. 2001, 290, 449–462. [Google Scholar] [CrossRef]
- Sember, A.; Nguyen, P.; Perez, M.F.; Altmanová, M.; Ráb, P.; Cioffi, M.B. Multiple sex chromosomes in teleost fishes from a cytogenetic perspective: State of the art and future challenges. Philos. Trans. R. Soc. B 2021, 376, 20200098. [Google Scholar] [CrossRef]
- Mariguela, T.C.; Roxo, F.F.; Foresti, F.; Oliveira, C. Phylogeny and biogeography of Triportheidae (Teleostei: Characiformes) based on molecular data. Mol. Phylogenetics Evol. 2016, 96, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Yano, C.F.; Sember, A.; Kretschmer, R.; Bertollo, L.A.C.; Ezaz, T.; Hatanaka, T.; Liehr, T.; Ráb, P.; Al-Rikabi, A.; Viana, P.F.; et al. Against the mainstream: Exceptional evolutionary stability of ZW sex chromosomes across the fish families Triportheidae and Gasteropelecidae (Teleostei: Characiformes). Chromosome Res. 2021, 29, 391–416. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Genera/Species by Family/Subfamily in Eschmeyer’s Catalog of Fishes. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 10 June 2023).
- Artoni, R.F.; Falcão, J.N.; Moreira-Filho, O.; Bertollo, L.A. An uncommon condition for a sex chromosome system in Characidae fish. Distribution and differentiation of ZZ/ZW system in Triportheus. Chromosome Res. 2001, 9, 449–456. [Google Scholar] [CrossRef]
- de Almeida Toledo, L.; Foresti, F. Morphologically differentiated sex chromosomes in neotropical freshwater fish. Genetica 2001, 111, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Cavallaro, Z.I. A highly differentiated ZZ/ZW sex chromosome system in a Characidae fish, Triportheus guentheri. Cytogenet. Cell Genet. 1992, 60, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Jorge, L.C. A new report of the ZZ/ZW sex chromosome system in the genus Triportheus. (Pisces, Triportheinae). Cytologia 1999, 64, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Artoni, R.F.; Bertollo, L.A.C. Evolutionary aspects of the ZZ/ZW sex chromosome system in the Characidae fish, genus Triportheus. A monophyletic state and NOR location on the W chromosome. Heredity 2002, 89, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of vertebrate sex chromosome evolution: From equal size to giants and dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Poltronieri, J.; Bertollo, L.A.C.; Artoni, R.F.; Liehr, T.; Cioffi, M.B. Chromosomal mapping of repetitive DNAs in Triportheus trifurcatus (Characidae, Characiformes): Insights into the differentiation of the Z and W chromosomes. PLoS ONE 2014, 9, e90946. [Google Scholar] [CrossRef]
- Nirchio, M.; Oliveira, C.; Ferreira, I.A.; Granado, A.; Ron, E. Extensive polymorphism and chromosomal characteristics of ribosomal DNA in the characid fish Triportheus venezuelensis (Characiformes, Characidae). Genet. Mol. Biol. 2007, 30, 25–30. [Google Scholar] [CrossRef]
- Diniz, D.; Laudicina, A.; Bertollo, L.A.C. Chromosomal location of 18S and 5S rDNA sites in Triportheus fish species (Characiformes, Characidae). Genet. Mol. Biol. 2009, 32, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Yano, C.F.; Bertollo, L.A.C.; Liehr, T.; Troy, W.P.; Cioffi, M.B. W chromosome dynamics in Triportheus species (Characiformes, Triportheidae)—An ongoing process narrated by repetitive sequences. J. Hered. 2016, 107, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.B.; Kejnovský, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The key role of repeated DNAs in sex chromosome evolution in two fish species with ZW sex chromosome system. Mol. Cytogenet. 2012, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Gimenez, O.M.; Dias, G.B.; De Lima, L.G.; Kuhn, G.C.E.S.; Ramos, É.; Martins, C.; Cabral-de-Mello, D.C. High-throughput analysis of the satellitome revealed enormous diversity of satellite DNAs in the neo-Y chromosome of the cricket Eneoptera surinamensis. Sci. Rep. 2017, 7, 6422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crepaldi, C.; Martí, E.; Gonçalves, É.M.; Martí, D.A.; Parise-Maltempi, P.P. Genomic Differences Between the Sexes in a Fish Species Seen Through Satellite DNAs. Front. Genet. 2021, 12, 728670. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lower, S.S.; McGurk, M.P.; Clark, A.G.; Barbash, D.A. Satellite DNA evolution: Old ideas, new approaches. Curr. Opin. Genet. Dev. 2018, 49, 70–78. [Google Scholar] [CrossRef]
- Ferretti, A.B.; Milani, D.; Palacios-Gimenez, O.M.; Ruiz-Ruano, F.J.; Cabral-de-Mello, D.C. High dynamism for neo-sex chromosomes: Satellite DNAs reveal complex evolution in a grasshopper. Heredity 2020, 125, 124–137. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Marec, F. Universal fluorescence in situ hybridization (FISH) protocol for mapping repetitive DNAs in insects and other arthropods. Mol. Genet. Genom. 2021, 296, 513–526. [Google Scholar] [CrossRef]
- Pereira, J.A.; Milani, D.; Ferretti, A.B.S.; Bardella, V.B.; Cabral-de-Mello, D.C.; Lopes, D.M. The extensive amplification of heterochromatin in Melipona bees revealed by high throughput genomic and chromosomal analysis. Chromosoma 2021, 130, 251–262. [Google Scholar] [CrossRef]
- de Lima, L.G.; Ruiz-Ruano, F.J. In-Depth Satellitome Analyses of 37 Drosophila Species Illuminate Repetitive DNA Evolution in the Drosophila Genus. Genome Biol. Evol. 2022, 14, evac064. [Google Scholar] [CrossRef]
- Montiel, E.E.; Mora, P.; Rico-Porras, J.M.; Palomeque, T.; Lorite, P. Satellitome of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), the most diverse among insects. Front. Ecol. Evol. 2022, 10, 826808. [Google Scholar] [CrossRef]
- Silva, D.M.D.A.; Utsunomia, R.; Ruiz-Ruano, F.J.; Daniel, S.N.; Porto-Foresti, F.; Hashimoto, D.T.; Foresti, F. High-throughput analysis unveils a highly shared satellite DNA library among three species of fish genus Astyanax. Sci. Rep. 2017, 7, 12726. [Google Scholar] [CrossRef] [Green Version]
- Utsunomia, R.; Silva, D.M.Z.D.A.; Ruiz-Ruano, F.J.; Goes, C.A.G.; Melo, S.; Ramos, L.P.; Hashimoto, D.T. Satellitome landscape analysis of Megaleporinus macrocephalus (Teleostei, Anostomidae) reveals intense accumulation of satellite sequences on the heteromorphic sex chromosome. Sci. Rep. 2019, 9, 5856. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Freitas, É.A.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Utsunomia, R.; Araya-Jaime, C.; Oliveira, C.; Camacho, J.P.M.; Foresti, F. Satellite DNA content of B chromosomes in the characid fish Characidium gomesi supports their origin from sex chromosomes. Mol. Genet. Genom. 2020, 295, 195–207. [Google Scholar] [CrossRef]
- Dos Santos, R.Z.; Calegari, R.M.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Melo, S.; Oliveira, C.; Foresti, F.; Uliano-Silva, M.; Porto-Foresti, F.; Utsunomia, R.A. Long-Term Conserved Satellite DNA That Remains Unexpanded in Several Genomes of Characiformes Fish Is Actively Transcribed. Genome Biol. Evol. 2021, 13, 2. [Google Scholar] [CrossRef]
- Kretschmer, R.; Goes, C.A.G.; Bertollo, L.A.C.; Ezaz, T.; Porto-Foresti, F.; Toma, G.A.; Utsunomia, R.; Cioffi, M.B. Satellitome analysis illuminates the evolution of ZW sex chromosomes of Triportheidae fishes (Teleostei: Characiformes). Chromosoma 2022, 131, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.J.; Destro, R.F.; Gazoni, T.; Narimatsu, H.; dos Santos, P.S.P.; Haddad, C.F.; Parise-Maltempi, P.P. Great abundance of satellite DNA in Proceratophrys (Anura, Odontophrynidae) revealed by genome sequencing. Cytogenet. Genome Res. 2020, 160, 141–147. [Google Scholar] [CrossRef]
- Silva, M.J.; Gazoni, T.; Haddad, C.F.B.; Parise-Maltempi, P.P. Analysis in Proceratophrys boiei genome illuminates the satellite DNA content in a frog from the Brazilian Atlantic forest. Front. Genet. 2023, 14, 1101397. [Google Scholar] [CrossRef] [PubMed]
- Lisachov, A.; Rumyantsev, A.; Prokopov, D.; Ferguson-Smith, M.; Trifonov, V. Conservation of Major Satellite DNAs in Snake Heterochromatin. Animals 2023, 13, 334. [Google Scholar] [CrossRef] [PubMed]
- Peona, V.; Kutschera, V.E.; Blom, M.P.; Irestedt, M.; Suh, A. Satellite DNA evolution in Corvoidea inferred from short and long reads. Mol. Ecol. 2023, 32, 1288–1305. [Google Scholar] [CrossRef] [PubMed]
- Valeri, M.P.; Dias, G.B.; do Espírito Santo, A.A.; Moreira, C.N.; Yonenaga-Yassuda, Y.; Sommer, I.B.; Svartman, M. First Description of a Satellite DNA in Manatees’ Centromeric Regions. Front. Genet. 2021, 12, 1537. [Google Scholar] [CrossRef]
- Sena, R.S.; Heringer, P.; Valeri, M.P.; Pereira, V.S.; Kuhn, G.C.S.; Svartman, M. Identification and characterization of satellite DNAs in two-toed sloths of the genus Choloepus (Megalonychidae, Xenarthra). Sci. Rep. 2020, 10, 19202. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Aleix-Mata, G.; Montiel, E.E.; Cabral-de-Mello, D.C.; Marchal, J.A.; Sánchez, A. Satellitome Analysis on Talpa aquitania Genome and Inferences about the satDNAs Evolution on Some Talpidae. Genes 2023, 14, 117. [Google Scholar] [CrossRef]
- Vozdova, M.; Kubickova, S.; Martínková, N.; Galindo, D.J.; Bernegossi, A.M.; Cernohorska, H.; Rubes, J. Satellite DNA in neotropical deer species. Genes 2021, 12, 123. [Google Scholar] [CrossRef] [PubMed]
- Šatović-Vukšić, E.; Plohl, M. Satellite DNAs—From Localized to Highly Dispersed Genome Components. Genes 2023, 14, 742. [Google Scholar] [CrossRef]
- Tyler-Smith, C.; Brown, W.R. Structure of the major block of alphoid satellite DNA on the human Y chromosome. J. Mol. Biol. 1987, 195, 457–470. [Google Scholar] [CrossRef]
- Moreira-Filho, O.; Bertollo, L.A.C. Extraction and use of the cephalic kidney for chromosome studies in small fish. Brazil J. Genet. 1991, 14, 1085–1090. [Google Scholar]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high sensitivity, Fluorescence hybridization. Proc. Nat. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Diniz, D.; Laudicina, A.; Cioffi, M.B.; Bertollo, L.A.C. Microdissection and whole chromosome painting. Improving sex chromosome analysis in Triportheus (Teleostei, Characiformes). Cytogenet. Genome Res. 2008, 122, 163–168. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Ezaz, T.; Trifonov, V.; Sember, A.; Liehr, T.; Cioffi, M.B. Highly conserved Z and molecularly diverged W chromosomes in the fish genus Triportheus (Characiformes, Triportheidae). Heredity 2017, 118, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, B.F.; Sidlauskas, B.L.; Near, T.J.; Roxo, F.F.; Ghezelayagh, A.; Ochoa, L.E.; Stiassny, M.L.J.; Arroyave, J.; Chang, J.; Faircloth, B.C.; et al. Accelerated diversification explains the exceptional species richness of tropical characoid fishes. Syst. Biol. 2021, 71, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Goes, C.A.G.; Santos, N.; Rodrigues, P.H.M.; Stornioli, J.H.F.; Silva, A.B.; Santos, R.Z.; Vidal, J.A.D.; Silva, D.M.Z.A.; Artoni, R.F.; Foresti, F.; et al. The Satellite DNA Catalogues of Two Serrasalmidae (Teleostei, Characiformes): Conservation of General satDNA Features over 30 Million Years. Genes 2023, 14, 91. [Google Scholar] [CrossRef]
- Rice, W.R. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef]
- Charlesworth, B. The evolution of chromosomal sex determination and dosage compensation. Curr. Biol. 1996, 6, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissensteiner, M.H.; Suh, A. Repetitive DNA: The dark matter of avian genomics. In Avian Genomics in Ecology and Evolution; Kraus, R., Ed.; Springer: Cham, Switzerland, 2019; pp. 93–150. [Google Scholar]
- McKee, B.D.; Handel, M.A. Sex chromosomes, recombination, and chromatin conformation. Chromosoma 1993, 102, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Gabriel-Robez, O.; Rumpler, Y.; Ratomponirina, C.; Petit, C.; Levilliers, J.; Croquette, M.F.; Couturier, J. Deletion of the pseudoautosomal region and lack of sex-chromosome pairing at pachytene in two infertile men carrying an X;Y translocation. Cytogenet. Cell Genet. 1990, 54, 38–42. [Google Scholar] [CrossRef]
- Cotter, D.J.; Brotman, S.M.; Sayres, M.A.W. Genetic diversity on the human X chromosome does not support a strict pseudoautosomal boundary. Genetics 2016, 203, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Page, D.C.; Bieker, K.; Brown, L.G.; Hinton, S.; Leppert, M.; Lalouel, J.M.; Lathrop, M.; Nystrom-Lahti, M.; de la Chapelle, A.; White, R. Linkage, physical mapping, and DNA sequence analysis of pseudoautosomal loci on the human X and Y chromosomes. Genomics 1987, 3, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, S.J.; Storer, J.M.; Hartley, G.A.; Grady, P.G.S.; Gershman, A.; de Lima, L.G.; Limouse, C.; Halabian, R.; Wojenski, L.; Rodriguez, M.; et al. From telomere to telomere: The transcriptional and epigenetic state of human repeat elements. Science 2022, 376, eabk3112. [Google Scholar] [CrossRef]
- Dover, G. Molecular drive: A cohesive mode of species evolution. Nature 1982, 299, 111–117. [Google Scholar] [CrossRef]
- Garrido-Ramos, M.A. Satellite DNA: An evolving topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Plohl, M.; Mestrovic, N.; Mravinac, B. Satellite DNA evolution. Genome Dyn. 2012, 7, 126–152. [Google Scholar] [CrossRef] [PubMed]
- Singchat, W.; O’Connor, R.E.; Tawichasri, P.; Suntronpong, A.; Sillpaprayoon, S.; Suntrarachun, S.; Muangmai, N.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; et al. Chromosome map of the Siamese cobra: Did partial synteny of sex chromosomes in the amniote represent “a hypothetical ancestral super-sex chromosome” or random distribution? BMC Genom. 2018, 19, 939. [Google Scholar] [CrossRef] [PubMed]
- Tomaszkiewicz, M.; Medvedev, P.; Makova, K.D. Y and W chromosome assemblies: Approaches and discoveries. Trends Genet. 2017, 33, 266–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.F.; Skaletsky, H.; Pyntikova, T.; Graves, T.A.; van Daalen, S.K.; Minx, P.J.; Fulton, R.S.; McGrath, S.D.; Locke, D.P.; Friedman, C.; et al. Chimpanzee and human Y chromosomes are remarkably divergent in structure and gene content. Nature 2010, 463, 536–539. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.A.M. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Yang, W.; An, N.; Zhang, P.; Zhang, G.; Zhou, Q. Temporal genomic evolution of bird sex chromosomes. BMC Evol. Biol. 2014, 14, 250. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; Nishida, C.; Matsuda, Y.; Kumazawa, Y. Sex chromosome evolution in snakes inferred from divergence patterns of two gametologous genes and chromosome distribution of sex chromosome-linked repetitive sequences. Zool. Lett. 2016, 2, 19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | N |
|---|---|---|
| Triportheus guentheri | São Francisco, MG | (12 ♀; 06 ♂) |
| Triportheus nematurus | Paraguai, MT | (09 ♀; 07 ♂) |
| Triportheus pantanensis | Paraguai, MT | (05 ♀; -- ♂) |
| Triportheus aff. rotundatus | Paraguai, MT | (19 ♀; 21 ♂) |
| Triportheus trifurcatus | Araguaia-Tocantins, MT | (04 ♀; 11 ♂) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, M.P.B.; Kretschmer, R.; Deon, G.A.; Toma, G.A.; Ezaz, T.; Goes, C.A.G.; Porto-Foresti, F.; Liehr, T.; Utsunomia, R.; Cioffi, M.d.B. Following the Pathway of W Chromosome Differentiation in Triportheus (Teleostei: Characiformes). Biology 2023, 12, 1114. https://doi.org/10.3390/biology12081114
de Oliveira MPB, Kretschmer R, Deon GA, Toma GA, Ezaz T, Goes CAG, Porto-Foresti F, Liehr T, Utsunomia R, Cioffi MdB. Following the Pathway of W Chromosome Differentiation in Triportheus (Teleostei: Characiformes). Biology. 2023; 12(8):1114. https://doi.org/10.3390/biology12081114
Chicago/Turabian Stylede Oliveira, Mariannah Pravatti Barcellos, Rafael Kretschmer, Geize Aparecida Deon, Gustavo Akira Toma, Tariq Ezaz, Caio Augusto Gomes Goes, Fábio Porto-Foresti, Thomas Liehr, Ricardo Utsunomia, and Marcelo de Bello Cioffi. 2023. "Following the Pathway of W Chromosome Differentiation in Triportheus (Teleostei: Characiformes)" Biology 12, no. 8: 1114. https://doi.org/10.3390/biology12081114