
Comparative Morphology of the Digestive Tract of African Bush Fish (Ctenopoma acutirostre) and Paradise Fish (Macropodus opercularis) Inhabiting Asian and African Freshwaters
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
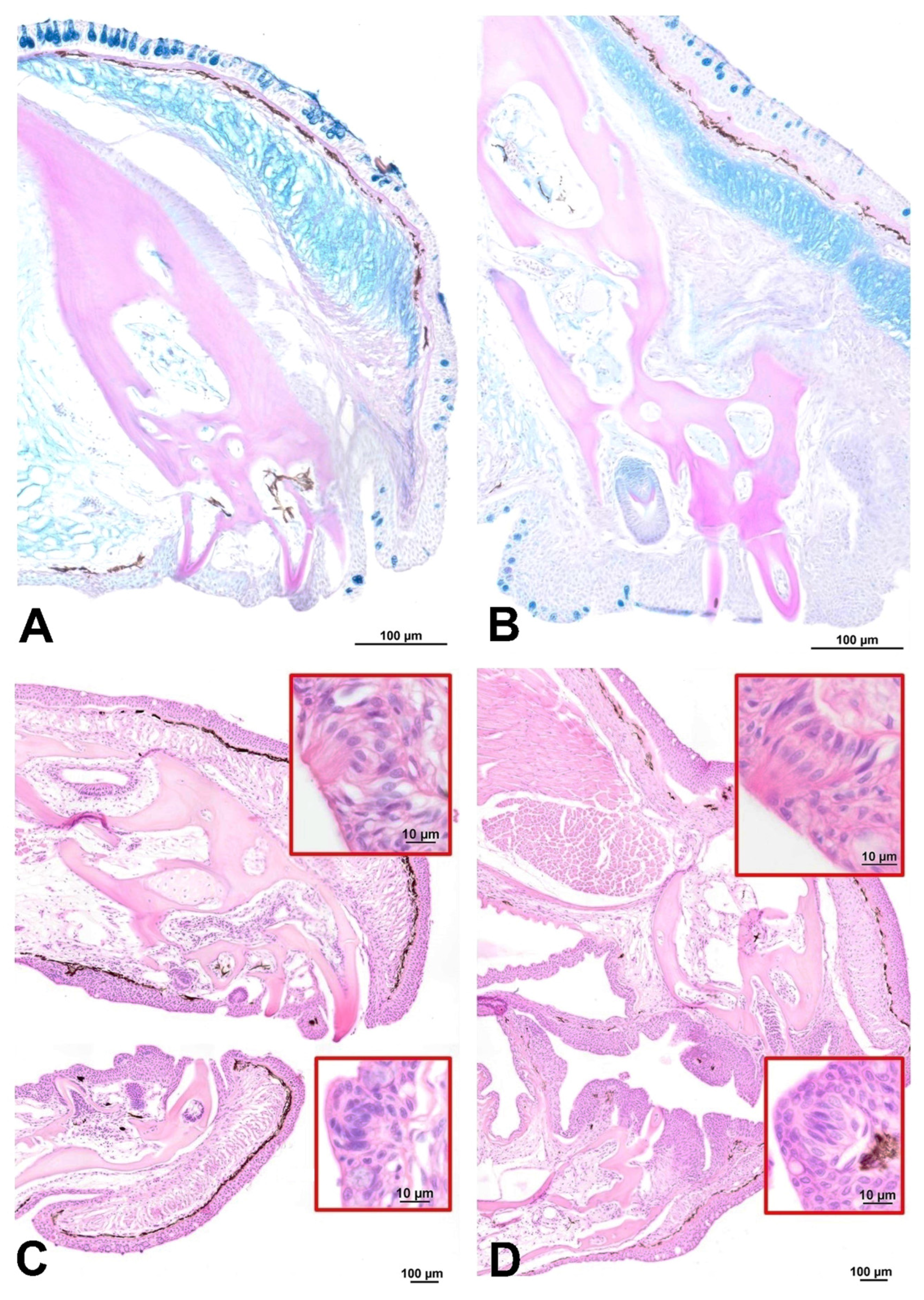
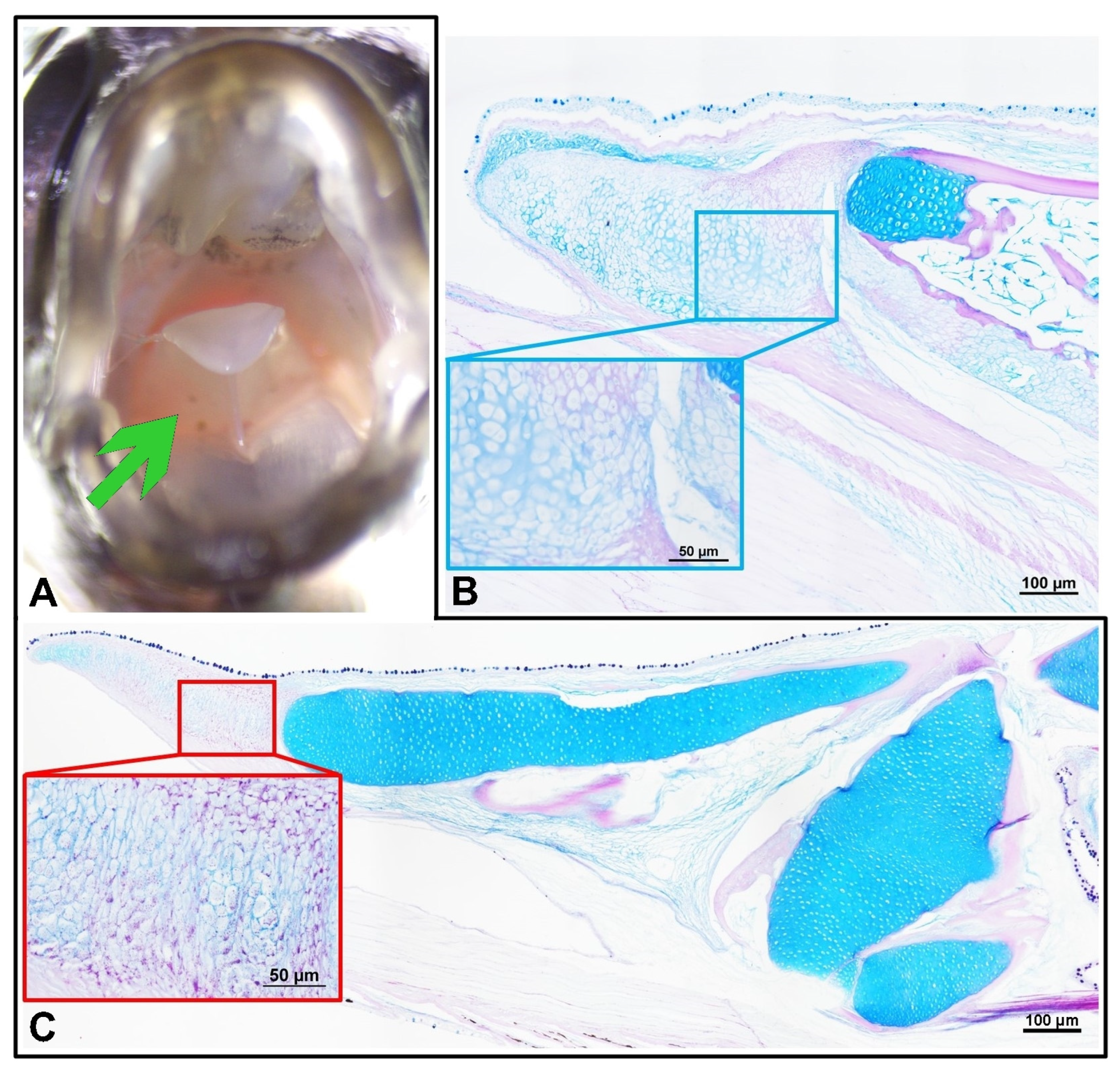
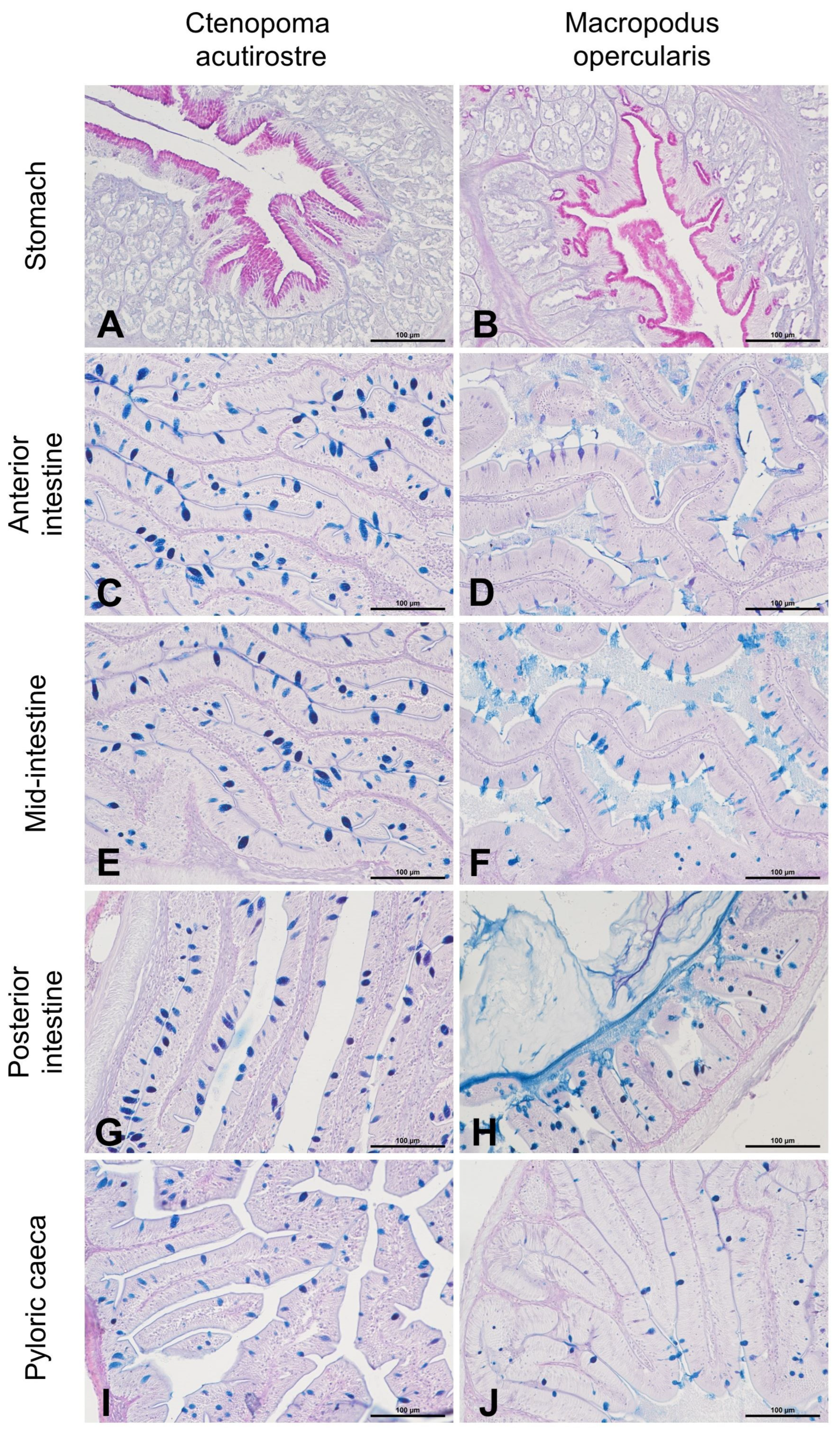
3. Results
Headgut
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, T.; Guo, Y. Early Normal Development of the Paradise Fish Macropodus Opercularis. Russ. J. Dev. Biol. 2018, 49, 240–244. [Google Scholar] [CrossRef]
- Evers, H.G.; Pinnegar, J.K.; Taylor, M.I. Where Are They All from?—Sources and Sustainability in the Ornamental Freshwater Fish Trade. J. Fish Biol. 2019, 94, 909–916. [Google Scholar] [CrossRef]
- Jones, M.; Alexander, M.E.; Snellgrove, D.; Smith, P.; Bramhall, S.; Carey, P.; Henriquez, F.L.; McLellan, I.; Sloman, K.A. How Should We Monitor Welfare in the Ornamental Fish Trade? Rev. Aquac. 2022, 14, 770–790. [Google Scholar] [CrossRef]
- Kowalska, A.; Kamaszewski, M.; Czarnowska-Kujawska, M.; Podlasz, P.; Kowalski, R.K. Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias Latipes). Animals 2020, 10, 2174. [Google Scholar] [CrossRef]
- Wiweger, M.; Majewski, L.; Adamek-Urbanska, D.; Wasilewska, I.; Kuznicki, J. Npc2-Deficient Zebrafish Reproduce Neurological and Inflammatory Symptoms of Niemann-Pick Type C Disease. Front. Cell. Neurosci. 2021, 15, 647860. [Google Scholar] [CrossRef]
- Reeves, G.A.; Singh, P.P.; Brunet, A. Chromatin Accessibility Profiling and Data Analysis Using ATAC-Seq in Nothobranchius Furzeri. In Cold Spring Harbor Protocols; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2023. [Google Scholar]
- Žák, J.; Anil, A.N.; Dyková, I. Dissolved Oxygen Saturation Is Crucial for Gas Bladder Inflation in Turquoise Killifish (Nothobranchius furzeri). Environ. Biol. Fishes 2023, 106, 673–683. [Google Scholar] [CrossRef]
- Rüber, L. Labyrinth Fishes (Anabantoidei). In The Timetree of Life; Oxford University Press: New Yourk, NY, USA, 2009; pp. 344–347. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2016; Volume 27, ISBN 9781119174844. [Google Scholar]
- Adamek-Urbańska, D.; Błażewicz, E.; Sobień, M.; Kasprzak, R.; Kamaszewski, M. Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes. Animals 2021, 11, 1158. [Google Scholar] [CrossRef]
- Dadgar, S.; Hafezieh, M.; Zargham, D.; Ghaedi, A.R. Optimum Dietary Protein Requirement of Paradise Fish, Macropodus Opercularis Based on Growth and Reproduction Performances. J. Surv. Fish. Sci. 2020, 7, 91–104. [Google Scholar] [CrossRef]
- Kitagawa, T.; Hosoya, K. Origin of the Ryukyuan Paradise Fish Macropodus Opercularis Inferred from Literature Survey. Biogeography 2016, 18, 11–16. [Google Scholar] [CrossRef]
- Miklosi, A.; Csanyi, V.; Gerlai, R. Antipredator Behavior in Paradise Fish (Macropodus Opercularis) Larvae: The Role of Genetic Factors and Paternal Influence. Behav. Genet. 1997, 27, 191–200. [Google Scholar] [CrossRef]
- Dutta, H.M. The Suspensorium of Ctenopoma Acutirostre: A Comparative Functional Analysis with Anabas Testudineus. J. Morphol. 1975, 146, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Prasetya, O.E.S.; Muarif, M.; Mumpuni, F.S. Pengaruh Pemberian pakan cacing sutera (Tubifex Sp.) Dan Daphnia Sp. Terhadap Pertumbuhan Dan Tingkat Kelangsungan Hidup Larva Ikan Lele Sangkuriang (Clarias gariepinus). J. Mina Sains 2020, 6, 8. [Google Scholar] [CrossRef]
- Santanumurti, M.B.; Nugroho, A.K.; Santoso, L.; Hudaidah, S. Study of Different Protein Content of Feeding of Local Raw Materials on Gourami Fish (Osphronemus Goramy Lac.) Aquaculture Performance. IOP Conf. Ser. Earth Environ. Sci. 2022, 1036, 012119. [Google Scholar] [CrossRef]
- Hickley, P.; Bailey, R.G. Food and Feeding Relationships of Fish in the Sudd Swamps (River Nile, Southern Sudan). J. Fish Biol. 1987, 30, 147–159. [Google Scholar] [CrossRef]
- Louca, V.; Lucas, M.C.; Green, C.; Majambere, S.; Fillinger, U.; Lindsay, S.W. Role of Fish as Predators of Mosquito Larvae on the Floodplain of the Gambia River. J. Med. Entomol. 2009, 46, 546–556. [Google Scholar] [CrossRef]
- Kassi, G.B.; Zeré, M.G.; Stanislas, S.Y.; N’douba, V. Régime Alimentaire de Ctenopoma Petherici (Perciformes, Anabantidae) Dans La Rivière Agnéby et Dans Le Lac de Barrage Hydroélectrique d’Ayamé 2 (Côte d’Ivoire). 2018. Available online: https://popups.uliege.be/2295-8010 (accessed on 13 June 2023).
- Rácz, A.; Adorján, G.; Fodor, E.; Sellyei, B.; Tolba, M.; Miklósi, Á.; Varga, M. Housing, Husbandry and Welfare of a “Classic” Fish Model, the Paradise Fish (Macropodus Opercularis). Animals 2021, 11, 786. [Google Scholar] [CrossRef]
- Puello-Cruz, A.C.; Velasco-Blanco, G.; Martínez-Rodríguez, I.E.; Felix-Ramos, E.; Voltolina, D. Growth and Survival of Siamese Fighting Fish, Betta Splendens,: Larvae at Low Salinity and with Different Diets. J. World Aquac. Soc. 2010, 41, 823–828. [Google Scholar] [CrossRef]
- Srikrishnan, R.; Hirimuthugoda, N.; Rajapakshe, W. Evaluation of Growth Performance and Breeding Habits of Fighting Fish (Betta Splendens) under 3 Diets and Shelters. J. Surv. Fish. Sci. 2017, 3, 50–65. [Google Scholar] [CrossRef]
- Pleeging, C.C.F.; Moons, C.P.H. Potential Welfare Issues of the Siamese Fighting Fish (Betta splendens) at the Retailer and in the Hobbyist Aquarium. Vlaams Diergeneeskd. Tijdschr. 2017, 86, 213–223. [Google Scholar] [CrossRef]
- Kilarski, W. Anatomia Ryb; Wydawnictwo Rolnicze i Leśne: Poznań, Poland, 2012; ISBN 9788309010807. [Google Scholar]
- Kasprzak, R.; Grzeszkiewicz, A.B.; Górecka, A. Performance of Co-Housed Neon Tetras (Paracheirodon innesi) and Glowlight Rasboras (Trigonostigma hengeli) Fed Commercial Flakes and Lyophilized Natural Food. Animals 2021, 11, 3520. [Google Scholar] [CrossRef]
- Ray, A.K.; Ringø, E. The Gastrointestinal Tract of Fish. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; Wiley: Hoboken, NJ, USA, 2014; pp. 1–13. [Google Scholar] [CrossRef]
- Karachle, P.K.; Stergiou, K.I. Intestine Morphometrics: A Compilation and Analysis of Bibliographic Data. Acta Ichthyol. Piscat. 2010, 40, 45–54. [Google Scholar] [CrossRef]
- German, D.P.; Horn, M.H. Gut Length and Mass in Herbivorous and Carnivorous Prickleback Fishes (Teleostei: Stichaeidae): Ontogenetic, Dietary, and Phylogenetic Effects. Mar. Biol. 2006, 148, 1123–1134. [Google Scholar] [CrossRef]
- Cho, J.H.; Park, J.W.; Ryu, Y.W.; Kim, K.W.; Hur, S.W. Morphology, Histology, and Histochemistry of the Digestive Tract of the Marbled Flounder Pseudopleuronectes Yokohamae. Animals 2023, 13, 936. [Google Scholar] [CrossRef] [PubMed]
- Karachle, P.K.; Stergiou, K.I. Gut Length for Several Marine Fish: Relationships with Body Length and Trophic Implications. Mar. Biodivers. Rec. 2010, 3, e106. [Google Scholar] [CrossRef]
- Omoniyi, I.T.; Agbon, A.O.; Akegbejo-Samsons, Y. The Food Habits of Ctenopoma pethereci, Gunther (Pisces: Anabantidae) in River Oluwa, Ondo State, Nigeria. West Afr. J. Appl. Ecol. 2012, 19, 1–7. [Google Scholar]
- Skelton, P.H. The Taxonomic Identity of Dwarf or Blackspot Ctenospoma (Pisces, Anabantidae) in Southern Africa. Cybium 1988, 12, 73–89. [Google Scholar]
- Buddington, R.K.; Kuz’mina, V. Digestive System. In The Laboratory Fish; Academic Press: Cambridge, MA, USA, 2000; pp. 379–384. [Google Scholar] [CrossRef]
- Meunier, F.J.; Brito, P.M.; Leal, M.-E.C. Morphological and Histological Data on the Structure of the Lingual Toothplate of Arapaima Gigas (Osteoglossidae; Teleostei). Cybium 2013, 37, 263–271. [Google Scholar]
- El Bakary, N.E.S.R. Morphological Study of the Asymmetrical Buccal Cavity of the Flatfish Common Solea (Solea Solea) and Its Relation to the Type of Feeding. Asian Pac. J. Trop. Biomed. 2014, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Meyer-rochow, V.B. Fish Tongues—Surface Fine Structures and Ecological Considerations. Zool. J. Linn. Soc. 1981, 71, 413–426. [Google Scholar] [CrossRef]
- Levanti, M.; Germanà, A.; Montalbano, G.; Guerrera, M.C.; Cavallaro, M.; Abbate, F. The Tongue Dorsal Surface in Fish: A Comparison Among Three Farmed Species. Anat. Histol. Embryol. 2017, 46, 103–109. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Ciriaco, E.; Germanà, A. Morphology of the Tongue Dorsal Surface of Gilthead Seabream (Sparus Aurata). Microsc. Res. Technol. 2012, 75, 1666–1671. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; De Carlos, F.; Suárez, A.Á.; Ciriaco, E.; Germanà, A. Morphology of the European Sea Bass (Dicentrarchus Labrax) Tongue. Microsc. Res. Technol. 2012, 75, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.L.; Brainerd, E.L. Role of Axial Muscles in Powering Mouth Expansion during Suction Feeding in Largemouth Bass (Micropterus salmoides). J. Exp. Biol. 2014, 217, 1333–1345. [Google Scholar]
- Sadeghinezhad, J.; Rahmati-Holasoo, H.; Fayyaz, S.; Zargar, A. Morphological Study of the Northern Pike (Esox lucius) Tongue. Anat. Sci. Int. 2015, 90, 235–239. [Google Scholar] [CrossRef]
- Palasai, A.; Lampang, P.N.; Gerald Plumley, F.; Kettratad, J.; Senarat, S.; Kangwanrangsan, N.; Jiraungkoorskul, W. Feeding Apparatus, Digestive System Structure, and Gut Contents of Priapium Fish, Neostethus Lankesteri Regan. Songklanakarin J. Sci. Technol. 1916, 44, 602–608. [Google Scholar]
- Le, H.T.M.D.; Shao, X.; Krogdahl, Å.; Kortner, T.M.; Lein, I.; Kousoulaki, K.; Lie, K.K.; Sæle, Ø. Intestinal Function of the Stomachless Fish, Ballan Wrasse (Labrus bergylta). Front. Mar. Sci. 2019, 6, 438019. [Google Scholar] [CrossRef]
- Hayakawa, Y. Influences of guarding territory on reproductive activity in the male dwarf gourami, colisa lalia. Indian J. Sci. Technol. 2011, 4, 113. [Google Scholar] [CrossRef]
- Nugraha, A.A.; Yustiati, A.; Bangkit, I.; Andriani, Y. Growth Performance and Survival Rate of Giant Gourami Fingerlings (Osphronemus Goramy Lacepede, 1801) with Potassium Diformate Addition. World Sci. News 2020, 143, 103–144. [Google Scholar]
- Boonanuntanasarn, S.; Jangprai, A.; Yoshizaki, G. Characterization of Proopiomelanocortin in the Snakeskin Gourami (Trichopodus pectoralis) and Its Expression in Relation to Food Intake. Domest. Anim. Endocrinol. 2015, 50, 1–13. [Google Scholar] [CrossRef]
- Tan, X.; Hoang, L.; Du, S.J. Characterization of Muscle-Regulatory Genes, Myf5 and Myogenin, from Striped Bass and Promoter Analysis of Muscle-Specific Expression. Mar. Biotechnol. 2002, 4, 537–545. [Google Scholar] [CrossRef]
- Moreira, E.; Novillo, M.; Eastman, J.T.; Barrera-Oro, E. Degree of Herbivory and Intestinal Morphology in Nine Notothenioid Fishes from the Western Antarctic Peninsula. Polar Biol. 2020, 43, 535–544. [Google Scholar] [CrossRef]
- Rust, M.B. Nutritional Physiology. In Fish Nutrition; Academic Press: Cambridge, MA, USA, 2003; pp. 367–452. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Length (cm) | Standard Length (cm) | Fork Length (cm) | Body Weight (g) | |
|---|---|---|---|---|
| Ctenopoma acutirostre | 5.39 ± 0.23 | 4.58 ± 0.24 | 0.82 ± 0.09 | 2.88 ± 0.56 |
| Macropodus opercularis | 5.47 ± 0.49 | 3.68 ± 0.18 | 1.79 ± 0.37 | 1.30 ± 0.49 |
| Length (cm) | Ctenopoma acutirostre | Macropodus opercularis |
|---|---|---|
| SL (cm) | 4.58 ± 0.24 | 3.68 ± 0.18 |
| IL (cm) | 3.38 ± 0.47 | 4.14 ± 0.70 |
| Pyloric caeca length (cm) | 0.25 ± 0.13 | 0.41 ± 0.09 |
| IL/SL | 0.74 * ± 0.09 | 1.13 * ± 0.14 |
| Range of IL/SL | 0.59–0.83 | 0.93–1.31 |
| % of body length | 59–83 | 93–131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamek-Urbańska, D.; Kamaszewski, M.; Wiechetek, W.; Wild, R.; Boczek, J.; Szczepański, A.; Śliwiński, J. Comparative Morphology of the Digestive Tract of African Bush Fish (Ctenopoma acutirostre) and Paradise Fish (Macropodus opercularis) Inhabiting Asian and African Freshwaters. Animals 2023, 13, 2613. https://doi.org/10.3390/ani13162613
Adamek-Urbańska D, Kamaszewski M, Wiechetek W, Wild R, Boczek J, Szczepański A, Śliwiński J. Comparative Morphology of the Digestive Tract of African Bush Fish (Ctenopoma acutirostre) and Paradise Fish (Macropodus opercularis) Inhabiting Asian and African Freshwaters. Animals. 2023; 13(16):2613. https://doi.org/10.3390/ani13162613
Chicago/Turabian StyleAdamek-Urbańska, Dobrochna, Maciej Kamaszewski, Wiktoria Wiechetek, Rafał Wild, Julia Boczek, Adrian Szczepański, and Jerzy Śliwiński. 2023. "Comparative Morphology of the Digestive Tract of African Bush Fish (Ctenopoma acutirostre) and Paradise Fish (Macropodus opercularis) Inhabiting Asian and African Freshwaters" Animals 13, no. 16: 2613. https://doi.org/10.3390/ani13162613