
Investigating the Ovarian Microstructure in the Genera Helicolenus and Scorpaena (Teleostei, Sub-Order Scorpaenoidei) with Implications for Ovarian Dynamics and Spawning

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Histological Preparation
2.3. Ovarian Dynamics
3. Results
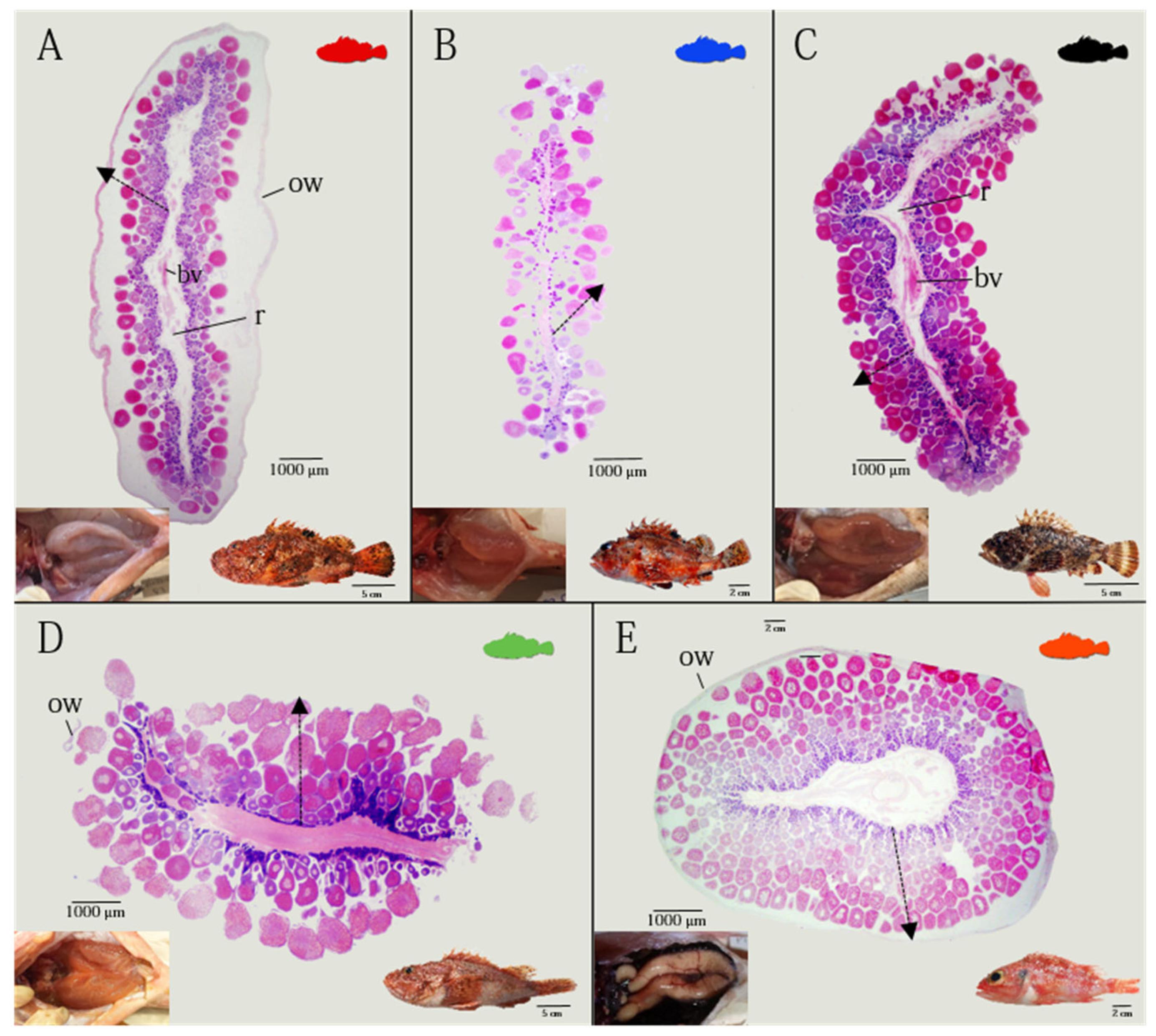
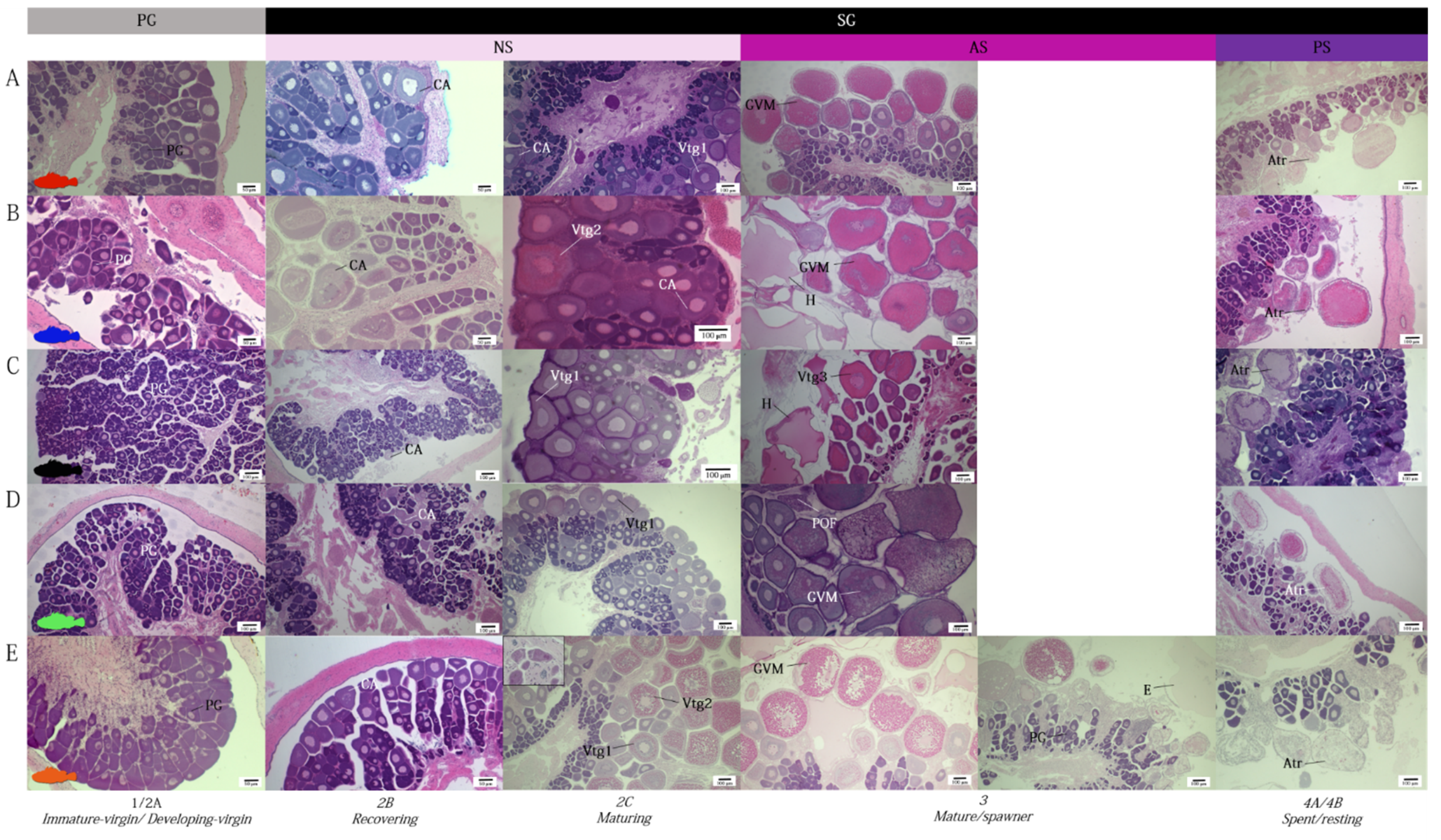
3.1. Ovarian Morphology
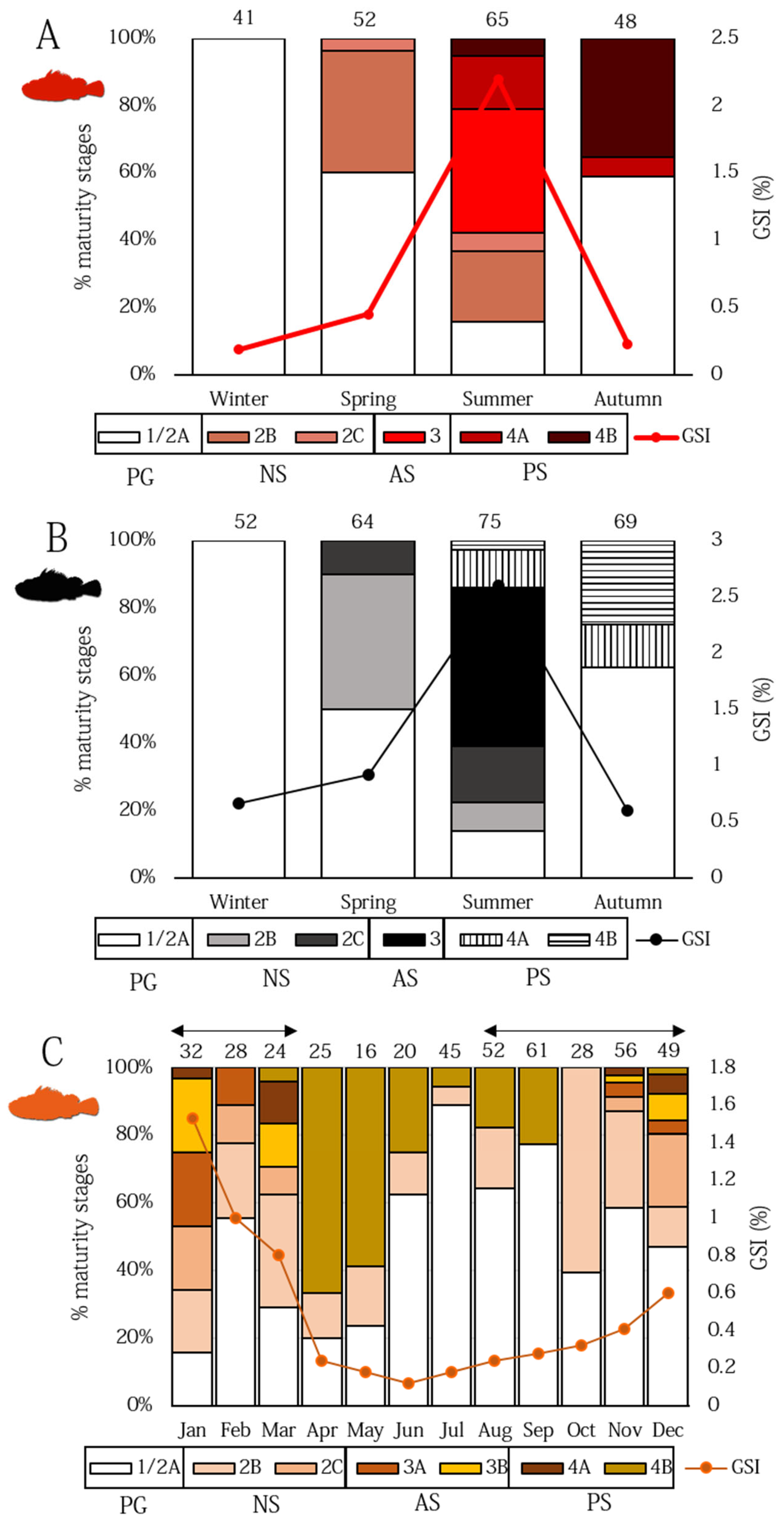
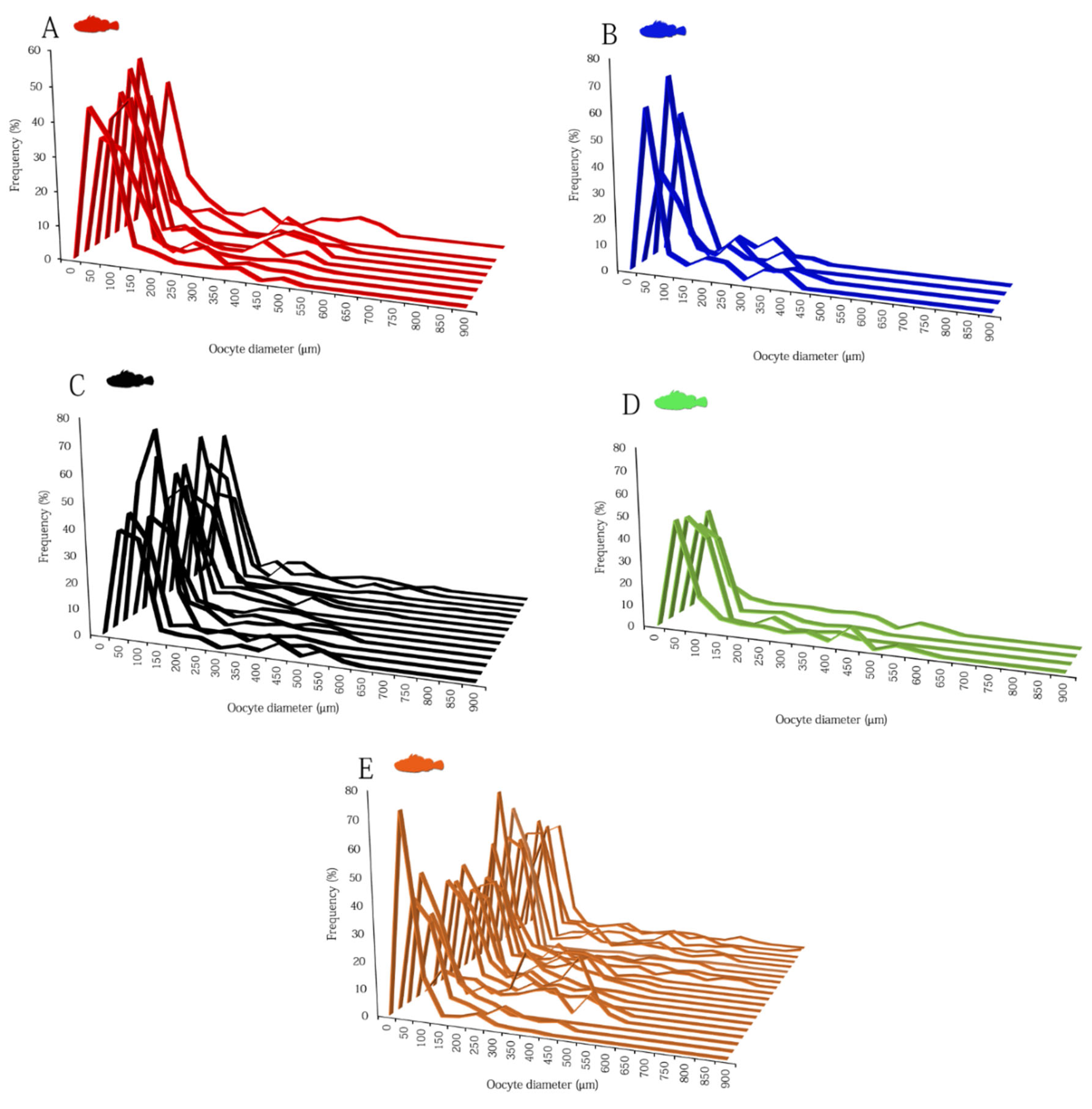
3.2. Ovarian Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wourms, J.P.; Lombardi, J. Reflections on the evolution of piscine viviparity. Am. Zool. 1992, 32, 276–293. [Google Scholar] [CrossRef]
- Wourms, J.P. Reproduction and development of Sebastes in the context of the evolution of piscine viviparity. Environ. Biol. Fish. 1991, 30, 111–126. [Google Scholar] [CrossRef]
- Muñoz, M. Reproduction in Scorpaeniformes. In Reproduction and Sexuality in Marine Fishes: Patterns and Processes; Cole, K.S., Ed.; University of California Press: Berkeley, CA, USA, 2010; Volume 3, pp. 65–90. [Google Scholar] [CrossRef]
- Blackburn, D.G.; Evans, H.E.; Vitt, L.J. The evolution of fetal nutritional adaptations. Fortschr Zool. 1985, 30, 437–439. [Google Scholar]
- Wourms, J.P.; Grove, B.D.; Lombardi, J. The maternal-embryonic relationship in viviparous fishes. Fish Physiol. 1988, 11, 1–134. [Google Scholar] [CrossRef]
- Koya, Y.; Muñoz, M. Comparative study on ovarian structures in scorpaenids: Possible evolutional process of reproductive mode. Ichthyol. Res. 2007, 54, 221–230. [Google Scholar] [CrossRef]
- Pavlov, D.A.; Emel’yanova, N.G. Transition to viviparity in the order Scorpaeniformes: Brief review. J. Ichthyol. 2013, 53, 52–69. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M.; Bonet, S. Annual reproductive cycle of Helicolenus dactylopterus dactylopterus (Teleostei: Scorpaeniformes) with special reference to the ovaries sperm storage. J. Mar. Biol. Assoc. 1999, 79, 521–529. [Google Scholar] [CrossRef]
- Ringvold, H.; Vesterinen, E.J. First in situ observations of the free-floating gelatinous matrix of blackbelly rosefish Helicolenus dactylopterus (Delaroche, 1809). Mar. Biol. Res. 2021, 17, 634–645. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M.; Bonet, S.; Quagio-Grassiotto, I. Sperm storage structures in the ovary of Helicolenus dactylopterus dactylopterus (Teleostei: Scorpaenidae): An ultrastructural study. Env. Biol. Fish. 2000, 58, 53–59. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M.; Bonet, S. The ovarian morphology of Scorpaena notata shows a specialized mode of oviparity. J. Fish Biol. 2002, 61, 877–887. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M.; Bonet, S. Testicular structure and semicystic spermatogenesis in a specialized ovuliparous species: Scorpaena notata (Pisces, Scorpaenidae). Acta Zool. 2002, 83, 213–219. [Google Scholar] [CrossRef]
- Vila, S. Annual Cycle of Stored Spermatozoa within the Ovaries of Helicolenus dactylopterus dactylopterus (Teleostei, Scorpaenidae). J. Fish Biol. 2007, 71, 596–609. [Google Scholar] [CrossRef]
- Sequeira, V.; Figueiredo, I.; Muñoz, M.; Gordo, L.S. New approach to the reproductive biology of Helicolenus dactylopterus. J. Fish Biol. 2003, 62, 1206–1210. [Google Scholar] [CrossRef]
- Sequeira, V.; Vila, S.; Neves, A.; Rifes, P.; Nunes, M.L.; Vieira, A.R.; Barros Paiva, R.; Muñoz, M.; Serrano Gordo, L. The gelatinous matrix of the Teleost Helicolenus dactylopterus dactylopterus (Delaroche, 1809) in the context of its reproductive strategy. Mar. Biol. Res. 2011, 7, 478–487. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.; Du, T.; Gao, G.; Wu, L.; Xu, S.; Xiao, Y.; Wang, Y.; Liu, Q.; Li, J. Sperm maturation, migration, and localization before and after copulation in black rockfish (Sebastes schlegelii). Theriogenology 2021, 166, 83–89. [Google Scholar] [CrossRef]
- Hoar, W.S. The gonads and reproduction. In Physiology of Fishes; Brown, M.E., Ed.; Academic Press: New York, NY, USA, 1957; pp. 287–321. [Google Scholar]
- Helfman, G.S.; Collette, B.B.; Facey, D.E.; Bowen, B.W. Teleosts at last I: Bonytongues through Anglerfishes. In The Diversity of Fishes: Biology, Evolution, and Ecology; Helfman, G.S., Collette, B.B., Facey, D.E., Bowen, B.W., Eds.; Blackwell: Boston, MA, USA, 2009; pp. 221–243. [Google Scholar]
- Takano, K. Ovarian structure and gametogenesis. In Reproductive Biology of Fish and Shellfish; Takashima, F., Hanyu, I., Eds.; Midori-shobo: Tokyo, Japan, 1989; pp. 33–34. [Google Scholar]
- Morris, J.A., Jr.; Sullivan, G.V.; Govoni, J.J. Oogenesis and spawn formation in the invasive lionfish, Pterois miles and Pterois volitans. Sci. Mar. 2011, 75, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Arculeo, M.; Lo Brutto, S. New contribution to the systematic status of various Mediterranean scorpionfish, as inferred from a mitochondrial DNA sequence. Rev. Biol. Mar. Oceanogr. 2014, 49, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. (Eds.) FishBase. Scorpaeniformes. In World Register of Marine Species; WoRMS: Ostend, Belgium, 2022; Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=10329 (accessed on 23 March 2022).
- Reglero, P.; Morales-Nin, B. Relationship between first sale price, body size and total catch of trammel net target species in Majorca (NW Mediterranean). Fish. Res. 2008, 92, 102–106. [Google Scholar] [CrossRef]
- Cadiou, G.; Boudouresque, C.F.; Bonhomme, P.; Le Diréach, L. The management of artisanal fishing within the Marine Protected Area of the Port-Cros National Park (Northwest Mediterranean Sea): A success story? ICES J. Mar. Sci. 2009, 66, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, P.; Romeo, T.; Consoli, P.; Scotti, G.; Andaloro, F. Characterization of the artisanal fishery and its socio-economic aspects in the central Mediterranean Sea (Aeolian Islands, Italy). Fish. Res. 2010, 102, 87–97. [Google Scholar] [CrossRef]
- Follesa, M.C.; Cannas, R.; Cau, A.; Cuccu, D.; Gastoni, A.; Ortu, A.; Pedoni, C.; Porcu, C.; Cau, A. Spillover effects of a Mediterranean marine protected area on the European spiny lobster Palinurus elephas (Fabricius, 1787) resource. Aquatic Conserv. Mar. Freshw. Ecosyst. 2011, 21, 564–572. [Google Scholar] [CrossRef]
- Ferri, J.; Stagličić, N.; Matić-Skoko, S. The black scorpionfish, Scorpaena porcus (Scorpaenidae): Could it serve as reliable indicator of Mediterranean coastal communities’ health? Ecol. Indic. 2012, 18, 25–30. [Google Scholar] [CrossRef]
- Muñoz, M.; Lloret, J.; Vila, S. Effects of artisanal fisheries on the scorpaenids (Scorpaena spp.) reproduction in the marine protected area of Cap de Creus (NW Mediterranean). Fish. Res. 2013, 138, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Follesa, M.C.; Porcu, C.; Cabiddu, S.; Mulas, A.; Deiana, A.M.; Cau, A. Deep-water fish assemblages in the central-western Mediterranean (south Sardinian deepwaters). J. Appl. Ichthyol. 2011, 27, 129–135. [Google Scholar] [CrossRef]
- Capezzuto, F.; Ancona, F.; Calculli, C.; Sion, L.; Maiorano, P.; D’Onghia, G. Feeding of the deep-water fish Helicolenus dactylopterus (Delaroche, 1809) in different habitats: From muddy bottoms to cold-water coral habitats. Deep-Sea Res. I 2020, 159, 103252. [Google Scholar] [CrossRef]
- Pirrera, L.; Bottari, T.; Busalacchi, B.; Giordano, D.; Modica, L.; Perdichizzi, A.; Profeta, A.; Rinelli, P. Distribution and population structure of the fish Helicolenus dactylopterus dactylopterus (Delaroche, 1809) in the central Mediterranean (Southern Tyrrhenian Sea). Mar. Ecol. 2009, 30, 161–174. [Google Scholar] [CrossRef]
- Sanchez, R.P.; Acha, E.M. Development and occurrence of embryos, larvae and juveniles of Sebastes oculafus with reference to two Southwest Atlantic scorpaenids: Helicolenus dactylopterus lahillei and Pontinus rathbuni. Meeresforsch 1988, 32, 107–133. [Google Scholar]
- White, D.B.; Wyanski, D.M.; Sedberry, G.R. Age, growth, and reproductive biology of the blackbelly rosefish from the Carolinas, U.S.A. J. Fish. Biol. 1998, 53, 1274–1291. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M. Reproductive indices and fecundity of Helicolenus dactylopterus dactylopterus (Teleostei: Scorpaenidae) in the Catalan Sea (Western Mediterranean). J. Mar. Biol. Assoc. UK 2002, 82, 995–1000. [Google Scholar] [CrossRef]
- Šegvić, T.; Grubišić, L.; Katavić, I.; Bartulović, V.; Pallaoro, A.; Dulčić, J. Embryonic and larval development of largescaled scorpionfish Scorpaena scrofa (Scorpaenidae). Cybium 2007, 31, 465–470. [Google Scholar]
- Shahrani, S.; Shakman, E. Biological aspects of Scorpaena scrofa Linnaeus, 1758 in the western Libyan coast. Libyan J. Mar. Sci. 2015, 14, 29–42. [Google Scholar]
- Sahin, C.; Erbay, M.; Kalayci, F.; Ceylan, Y.; Yesilcicek, T. Life-history traits of the black scorpionfish (Scorpaena porcus) in SouthEastern Black Sea. Turk. J. Fish. Aquat. Sci. 2019, 19, 571–584. [Google Scholar] [CrossRef]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Follesa, M.C.; Agus, B.; Bellodi, A.; Cannas, R.; Capezzuto, F.; Casciaro, L.; Cau, A.; Cuccu, D.; Donnaloia, M.; Fernandez-Arcaya, U. The MEDITS maturity scales as a useful tool for investigating the reproductive traits of key species in the Mediterranean Sea. Sci. Mar. 2019, 83, 235–256. [Google Scholar] [CrossRef] [Green Version]
- Follesa, M.C.; Carbonara, P. Atlas of the maturity stages of Mediterranean fishery resources. Gen. Fish. Comm. Mediterr. Stud. Rev. 2019, 99, 1–259. [Google Scholar]
- Cerri, P.S.; Sasso-Cerri, E. Staining methods applied to glycol methacrylate embedded tissue sections. Micron 2003, 34, 365–372. [Google Scholar] [CrossRef]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Hunter, J.R.; Macewicz, B.J. Measurement of spawning frequency in multiple spawning fishes. NOAA Tech. Rep. NMFS 1985, 36, 79–94. [Google Scholar]
- Rohlf, F.J. TpsDig, Digitize Landmarks and Outlines, Version 2.16. In Department of Ecology and Evolution; State University of New York at Stony Brook: New York, NY, USA, 2005. [Google Scholar]
- West, G.J. Methods of assessing ovarian development inn fishes: A review. Aust. J. Mar. Freshw. Res. 1990, 41, 199–222. [Google Scholar] [CrossRef]
- Cooper, D.W.; Maslenikov, K.P.; Gunderson, D.R.; Gunderson, D.R. Natural mortality rate, annual fecundity, and maturity at length for Greenland halibut (Reinhardtius hippoglossoides) from the northeastern Pacific Ocean. Fish. Bull. 2007, 105, 296–304. [Google Scholar]
- Carbonara, P.; Porcu, C.; Donnaloia, M.; Pesci, P.; Sion, L.; Spedicato, M.T.; Zupa, W.; Vitale, F.; Follesa, M.C. The spawning strategy of European hake (Merluccius merluccius, L. 1758) across the western and central Mediterranean Sea. Fish. Res. 2019, 219, 105333. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 663. [Google Scholar]
- Karlou-Riga, C.; Petza, D.; Charitonidou, K.; Anastopoulos, P.; Koulmpaloglou, D.S.; Ganias, K. Ovarian dynamics in picarel (Spicara smaris, L., Sparidae) and implications for batch fecundity and spawning interval estimation. J. Sea Res. 2020, 160, 101894. [Google Scholar] [CrossRef]
- Hoar, W.S. 1. Reproduction. In Physiology of Fishes; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA, 1969; Volume 3, pp. 1–72. [Google Scholar]
- Brummett, A.R.; Dumont, J.N.; Larkin, J.R. The ovary of Fundulus heteroclitus. J. Morph. 1982, 173, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.L. The structure of the ovary-vertebrates. In The Ovary; Zuckerman, S., Ed.; Academic Press: London, UK, 1962; Volume 1. [Google Scholar]
- Fishelson, L. Ethology and reproduction of pteroid fishes found in the Gulf of Agaba (Red Sea), especially Dendrochirus brachypterus (Cuvier), (Pteroidae, Teleostei). Pubbl. Staz. Zool. Napoli 1975, 39, 635–656. [Google Scholar]
- Nagahama, Y. The functional morphology of teleost gonads. In Fish Physiology; Hoar, W.S., Randall, D.J., Donaldson, E.M., Eds.; Academic Press: New York, NY, USA, 1983; Volume 9, pp. 223–275. [Google Scholar]
- Orton, G.L. Early developmental stages of the California scorpionfish, Scorpaena guttata. Copeia 1955, 3, 210–214. [Google Scholar] [CrossRef]
- Washington, B.B.; Moser, H.G.; Laroche, W.A.; Richards, W.J., Jr. Scorpaeniformes: Development. In Ontogeny and Systematics of Fishes; Moser, H.G., Richards, W.J., Cohen, D.M., Fahay, M.P., Kendall, A.W., Jr., Richardson, S.L., Eds.; American Society of Ichthyologists and Herpetologists Special Publication Number 1; Allen Press: Lawrence, KS, USA, 1984; pp. 405–428. [Google Scholar]
- Pavlov, D.A.; Emel’yanova, N.G. Features of reproductive biology in two tropical fish species from the family Scorpaenidae. J. Ichthyol. 2007, 47, 353–365. [Google Scholar] [CrossRef]
- Emel’yanova, N.G.; Pavlov, D.A. Features of reproductive biology of Papuan scorpionfish Scorpaenopsis papuensis (Scorpaenidae). J. Ichthyol. 2021, 61, 143–151. [Google Scholar] [CrossRef]
- Riehl, R.; Greven, H. Fine structure of egg envelopes in some viviparous goodeid fishes, with comments on the relation of envelope thinness to viviparity. Can. J. Zool. 1993, 71, 91–97. [Google Scholar] [CrossRef]
- Follesa, M.C.; Porcu, C.; Mulas, A.; Salvadori, S.; Cau, A. Reproductive characteristics of the bathyal viviparous fish Cataetyx alleni (Osteichthyes: Bythitidae) from the southeastern Sardinian Sea (central-western Mediterranean). Sci. Mar. 2011, 75, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Tengfei, D.; Yongshuang, X.; Haixia, Z.; Li, Z.; Qinghua, L.; Xueying, W.; Jun, L.; Shihong, X.; Yanfeng, W.; Jiachen, Y.; et al. Multiple fetal nutritional patterns before parturition in viviparous fish Sebastes schlegelii (Hilgendorf, 1880). Front. Mar. Sci. 2021, 7, 571946. [Google Scholar] [CrossRef]
- Xiaojie, X.; Xueying, W.; Qinghua, L.; Xin, Q.; Li, Z.; Haoming, L.; Jun, L. New insights on folliculogenesis and follicular placentation in marine viviparous fish black rockfish (Sebastes schlegelii). Gene 2022, 827, 146444. [Google Scholar] [CrossRef]
- Muñoz, M.; Casadevall, M.; Bonet, S. Gonadal structure and gametogenesis of Aspitrigla obscura (Pisces, Triglidae). Ital. J. Zool. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Pavlov, D.A.; Emel’yanova, N.G. Biological characteristics of Dendrochirus zebra (Cuvier, 1829) (Scorpaeniformes: Scorpaenidae) from Nha Trang Bay, South China Sea. Russ. J. Mar. Biol. 2019, 45, 75–85. [Google Scholar] [CrossRef]
- Selman, K.; Wallace, R.A. Cellular aspects of oocyte growth in teleosts. Zool. Sci. 1989, 6, 211–231. [Google Scholar]
- Rizzo, E.; Sato, Y.; Barreto, B.P.; Godinho, H.P. Adhesiveness and surface patterns of eggs in neotropical freshwater teleosts. J. Fish Biol. 2005, 61, 615–632. [Google Scholar] [CrossRef]
- Nakashima, S.; Iwamatsu, T. Ultrastructural changes in micropylar and granulosa cells during in vitro oocyte maturation in the medaka, Oryzias latipes. J. Exp. Zool. 1994, 270, 547–556. [Google Scholar] [CrossRef]
- Żelazowska, M. Formation and structure of egg envelopes in Russian sturgeon Acipenser gueldenstaedtii (Acipenseriformes: Acipenseridae). J. Fish Biol. 2010, 76, 694–706. [Google Scholar] [CrossRef]
- Moser, H.G. Reproduction and development of Sebastodes paucispinis and comparison with other rockfishes off southern California. Copeia 1967, 1967, 773–797. [Google Scholar] [CrossRef]
- Koya, Y.; Munehara, H.; Takano, K. Sperm storage and degradation in the ovary of a marine copulating sculpin, Alcichthys alcicornis (Teleostei: Scorpaeniformes): Role of intercellular junctions between inner ovarian epithelial cells. J. Morph. 1997, 233, 153–163. [Google Scholar] [CrossRef]
- Bradai, M.N.; Bouain, A. Reproduction de Scorpaena porcus et de Scorpena scrofa (L., 1758) (Pisces, Scorpaenidae) du Golfe de Gabes. Oebalia 1991, 17, 167–180. [Google Scholar]
- Koca, H.U. Sinop yöresinde dip ağlarla ile avlanan iskorpit (Scorpaena porcus Linne., 1758) balığının balıkçılık biyolojisi yönünden bazı özelliklerinin araştırılması. Turk. J. Vet. Anim. Sci. 2002, 26, 65–69. [Google Scholar]
- Muñoz, M.; Sàbat, M.; Vila, S.; Casadevall, M. Annual reproductive cycle and fecundity of Scorpaena notata (Teleostei, Scorpaenidae). Sci. Mar. 2005, 69, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, A.; Isidro, E.; Menezes, G.; Pinho, M.R.; Melo, O.; Estácio, S. New contribution to the reproductive features of bluemouth Helicolenus dactylopterus dactylopterus from northeast Atlantic (Azores Archipelago). Sci. Mar. 2006, 70, 679–688. [Google Scholar] [CrossRef]
- Sequeira, V.; Neves, A.; Paiva, R.B.; de Lima, J.P.; Vieira, A.R.; Gordo, L.S. Life history parameters as possible discriminators of bluemouth Helicolenus dactylopterus (Delaroche, 1809) populations in Portuguese waters. Fish. Res. 2012, 125, 69–76. [Google Scholar] [CrossRef]
- Pavlov, D.A.; Emel’yanova, N.G. Reproductive biology of species from the family Scorpaenidae and transition from oviparity to viviparity in the southern and northern Percomorpha. In Viviparous Fishes II; Uribe, M.C., Grier, H.J., Eds.; New Life: Homestead, FL, USA, 2010; pp. 89–105. [Google Scholar]
- Muñoz, M.; Dimitriadis, C.; Casadevall, M.; Vila, S.; Delgado, E.; Lloret, J.; Saborido-Rey, F. Female reproductive biology of the bluemouth Helicolenus dactylopterus dactylopterus: Spawning and fecundity. J. Fish Biol. 2010, 77, 2423–2442. [Google Scholar] [CrossRef]
- Ganias, K.; Lowerre-Barbieri, S. Oocyte recruitment and fecundity type in fishes: Refining terms to reflect underlying processes and drivers. Fish Fish. 2018, 19, 562–572. [Google Scholar] [CrossRef]
- Hunter, J.R.; Lo, N.C.H.; Moser, H.G.; Smith, P.E.; Methot, R.D. The daily fecundity reduction method: A new procedure for estimating adult fish biomass. ICES J. Mar. Sci. 1992, 49, 209–215. [Google Scholar]
- Neves, A.; Vieira, A.R.; Farias, I.; Figueiredo, I.; Sequeira, V.; Gordo, L.S. Reproductive strategies in black scabbardfish (Aphanopus carbo Lowe, 1839) from the NE Atlantic. Sci. Mar. 2009, 73, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Jansen, T.; Slotte, A.; dos Santos Schmidt, T.C.; Sparrevohn, C.R.; Jacobsen, J.A.; Kjesbu, O.S. Bioenergetics of egg production in northeast Atlantic mackerel changes the perception of fecundity type and annual trends in spawning stock biomass. Progr. Oceanogr. 2021, 198, 102658. [Google Scholar] [CrossRef]
- Dos Santos Schmidt, T.C.; Thorsen, A.; Slotte, A.; Nøttestad, L.; Kjesbu, O.S. First thorough assessment of de novo oocyte recruitment in a teleost serial spawner, the Northeast Atlantic mackerel (Scomber scombrus) case. Sci. Rep. 2021, 11, 21795. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Size Range (TL, cm) | Depth Range (m) |
|---|---|---|---|
| Family Scorpaenidae | |||
| Scorpaena elongata Cadenat, 1943 | 45 | 19.5–47.5 | 60–478 |
| Scorpaena notata Rafinesque, 1810 | 33 | 8.1–14.2 | 13–101 |
| Scorpaena porcus Linnaeus, 1758 | 260 | 9.1–21.6 | 21–155 |
| Scorpaena scrofa Linnaeus, 1758 | 206 | 9.5–36.9 | 30–155 |
| Family Sebastidae | |||
| Helicolenus dactylopterus (Delaroche, 1809) | 436 | 7.8–34.2 | 60–651 |
| Structures | PAS | AB pH 2.5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HD | SE | SN | SP | SS | HD | SE | SN | SP | SS | |
| Ovarian wall | + | + | + | + | + | + | + | + | + | + |
| Ovarian fluid | + | + | + | + | + | − | − | − | − | − |
| Holocrine gland | + | + | + | + | + | − | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porcu, C.; Lai, E.; Bellodi, A.; Carbonara, P.; Cau, A.; Mulas, A.; Pascale, N.; Porceddu, R.; Follesa, M.C. Investigating the Ovarian Microstructure in the Genera Helicolenus and Scorpaena (Teleostei, Sub-Order Scorpaenoidei) with Implications for Ovarian Dynamics and Spawning. Animals 2022, 12, 1412. https://doi.org/10.3390/ani12111412
Porcu C, Lai E, Bellodi A, Carbonara P, Cau A, Mulas A, Pascale N, Porceddu R, Follesa MC. Investigating the Ovarian Microstructure in the Genera Helicolenus and Scorpaena (Teleostei, Sub-Order Scorpaenoidei) with Implications for Ovarian Dynamics and Spawning. Animals. 2022; 12(11):1412. https://doi.org/10.3390/ani12111412
Chicago/Turabian StylePorcu, Cristina, Eleonora Lai, Andrea Bellodi, Pierluigi Carbonara, Alessandro Cau, Antonello Mulas, Noemi Pascale, Riccardo Porceddu, and Maria Cristina Follesa. 2022. "Investigating the Ovarian Microstructure in the Genera Helicolenus and Scorpaena (Teleostei, Sub-Order Scorpaenoidei) with Implications for Ovarian Dynamics and Spawning" Animals 12, no. 11: 1412. https://doi.org/10.3390/ani12111412