Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Diets
- Vegetable waste (mixed peels of 10% onion, 25% potato, 25% sweet potato, 30% carrot, and 10% cucumber, with a total water content of 91.4%);
- Green garden waste with grass (50% Poaceae species and other common weeds, 25% tree leaves, and 25% branches (Populus, Salix, Pinus, and Corylus species), and a mixture of stone fruits, and other ornamental plant parts, with a water content of 36.2%;
- 55% cattle manure with faeces and urine, and 45% cereal straw with a water content of 45.7%;
- 35% horse manure with faeces and urine, and 65% cereal straw with a water content of 28.3%.
2.2. Experimental Design
2.3. Measured Parameters
2.4. Data Analyses
3. Results
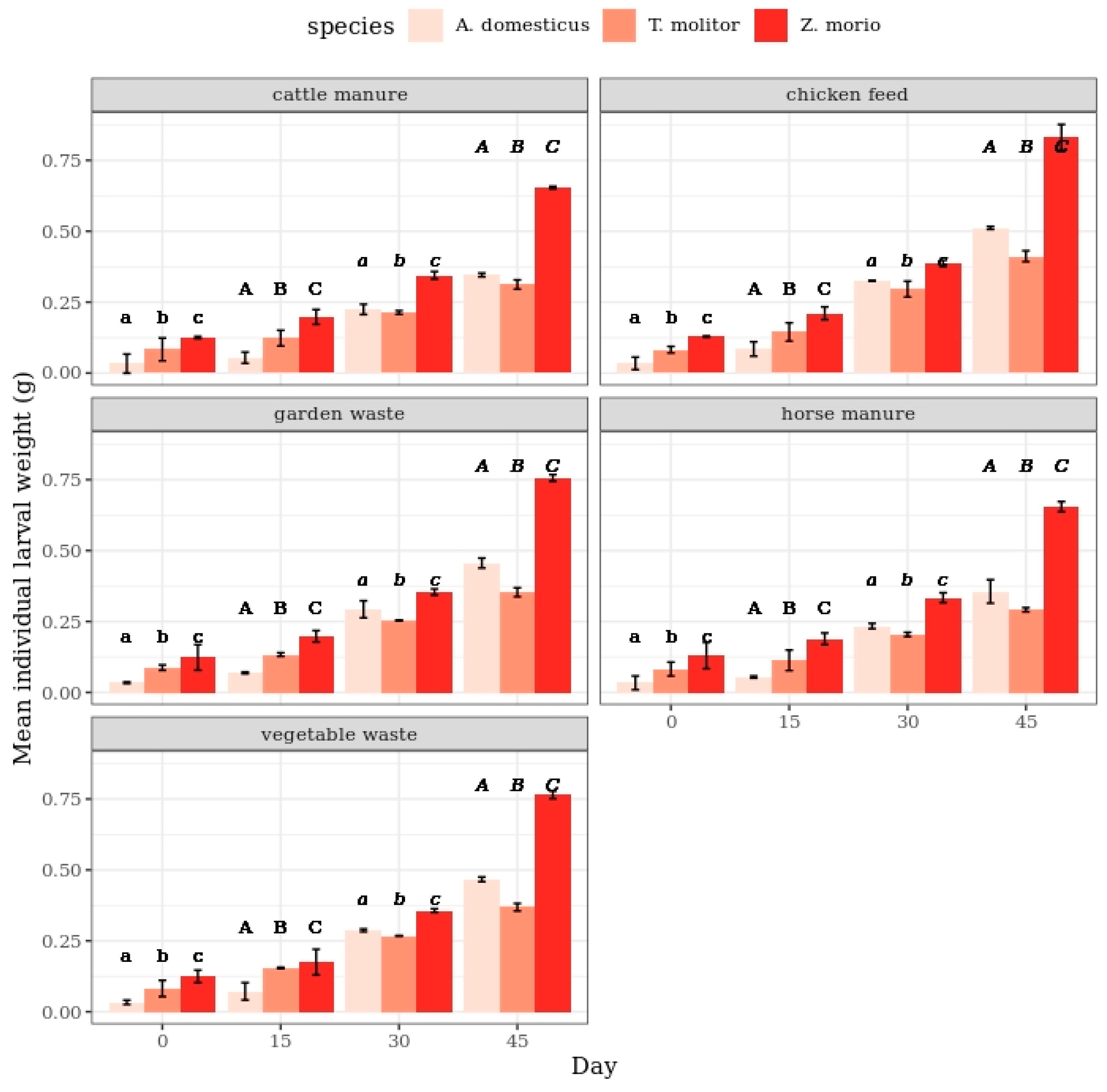
3.1. Effect of Substrates on Larval Weight
3.2. Nutritional Compositions of the Substrates
3.3. Nutrient Value of the Larvae
3.4. Evaluation of the Effect of Substrates and Rearing Time on the Survival of Larvae
4. Discussion
4.1. Changes in the Larval Weight in Response to Diet
4.2. Nutritional Value of the Larvae
4.3. Mass of Live Larvae during Rearing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Benzertiha, A.; Kieronczyk, B.; Kołodziejski, P.; Pruszynska–Oszmałek, E.; Rawski, M.; Józefiak, D.; Józefiak, A. Tenebrio molitor and Zophobas morio full-fat meals as functional feed additives affect broiler chickens’ growth performance and immune system traits. Poult. Sci. 2020, 99, 196–206. [Google Scholar] [CrossRef]
- De Goede, D.M.; Erens, J.; Kapsomenou, E.; Peters, M. Large scale insect rearing and animal welfare. In The Ethics of Consumption; Röcklinsberg, H., Sandin, P., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2013; pp. 236–242. ISBN 9789086862313. [Google Scholar]
- Ortiz, J.A.C.; Ruiz, A.T.; Morales-Ramos, J.A.; Thomas, M.; Rojas, M.G.; Tomberlin, J.K.; Yi, L.; Han, R.; Giroud, L.; Jullien, R.L. Insect mass production technologies. In Insects as Sustainable Food Ingredients: Production, Processing, and Food Applications; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: London, UK, 2016; pp. 153–201. ISBN 9780128028568. [Google Scholar]
- Chuck-Hernández, C.; Allende, D.K.B.; Mahlknecht, J. Novel food ingredients for food security. Encycl. Food Secur. Sustain. 2019, 1, 369–375. [Google Scholar] [CrossRef]
- Varelas, V. Food wastes as a potential new source for edible insect mass production for food and feed: A review. Fermentation 2019, 5, 81. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumo, M.; Osuga, I.M.; Khamis, F.M.; Tanga, C.M.; Fiaboe, K.K.M.; Subramanian, S.; Ekesi, S.; van Huis, A.; Borgemeister, C. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 2019, 9, 10110. [Google Scholar] [CrossRef]
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect rearing: Potential, challenges, and circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Dossey, A.T. Age-dependent food utilisation of Acheta domesticus (Orthoptera: Gryllidae) in small groups at two temperatures. J. Insects Food Feed 2018, 4, 51–60. [Google Scholar] [CrossRef]
- Lähteenmäki-Uutela, A.; Grmelová, N. European law on insects in food and feed. Eur. Food Feed Law Rev. 2016, 11, 2–8. [Google Scholar] [CrossRef]
- Skrivervik, E. Insects’ contribution to the bioeconomy and the reduction of food waste. Heliyon 2020, 6, e03934. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Tomberlin, J.K.; Vanlaerhoven, S. Influence of resources on Hermetia illucens (Diptera: Stratiomyidae) larval development. J. Med. Entomol. 2013, 50, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Varelas, V.; Langton, M. Forest biomass waste as a potential innovative source for rearing edible insects for food and feed—A review. Innov. Food Sci. Emerg. Technol. 2017, 41, 193–205. [Google Scholar] [CrossRef]
- Kim, S.Y.; Chung, T.-H.; Kim, S.-H.; Song, S.; Kim, N. Recycling agricultural wastes as feed for mealworm (Tenebrio molitor). Korean J. Appl. Entomol. 2014, 53, 367–373. [Google Scholar] [CrossRef]
- Nyakeri, E.M.; Ogola, H.J.O.; Ayieko, M.A.; Amimo, F.A. Valorisation of organic waste material: Growth performance of wild black soldier fly larvae (Hermetia illucens) reared on different organic wastes. J. Insects as Food Feed 2017, 3, 193–202. [Google Scholar] [CrossRef]
- Zaelor, J.; Kitthawee, S. Growth response to population density in larval stage of darkling beetles (Coleoptera; Tenebrionidae) Tenebrio molitor and Zophobas atratus. Agric. Nat. Resour. 2018, 52, 603–606. [Google Scholar] [CrossRef]
- Lundy, M.E.; Parrella, M.P. Crickets are not a free lunch: Protein capture from scalable organic side-streams via high-density populations of Acheta domesticus. PLoS ONE 2015, 10, e0118785. [Google Scholar] [CrossRef]
- Jucker, C.; Lupi, D.; Moore, C.D.; Leonardi, M.G.; Savoldelli, S. Nutrient Recapture from Insect Farm Waste: Bioconversion with Hermetia illucens (L.) (Diptera: Stratiomyidae). Sustainability 2020, 12, 362. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth performance and nutrient composition of mealworms (Tenebrio Molitor) fed on fresh plant materials—Supplemented diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Adámková, A.; Adámek, M.; Mlček, J.; Borkovcová, M.; Bednářová, M.; Kouřimská, L.; Skácel, J.; Vítová, E. Welfare of the mealworm (Tenebrio molitor) breeding with regard to nutrition value and food safety. Potravin. Slovak J. Food Sci. 2017, 11, 460–465. [Google Scholar]
- Booth, D.T.; Kiddell, K. Temperature and energetics of development in the house cricket (Acheta domesticus). J. Insect Physiol. 2007, 53, 950–953. [Google Scholar] [CrossRef]
- Miech, P.; Berggen, A.; Lindberg, J.E.; Chhay, T.; Khieu, B.; Jansson, A. Growth and survival or reared Cambodian field crickets (Releogryllus testaceus) fed weeds, agricultural and food industry by-products. J. Insects Food Feed 2016, 2, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.L.; Berggreen, I.E.; Heckmann, L.-H.L. Recommendations for Breeding and Holding of Regular Mealworm, Tenebrio Molitor; Insect Group, Water and Environment, Institute of Technology: Aarhus, Denmark, 2017; Available online: https://www.bugburger.se/wp-content/uploads/2018/11/mealwormguide.pdf (accessed on 1 July 2020).
- Coudron, C.; Spranghers, T.; Elliot, D.; Halstead, J. Report on the Project Entitled BioBoost—Insect Breeding. 2019. Available online: https://www.BioBoosteurope.com (accessed on 1 July 2020).
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Adámková, A.; Mlcek, J.; Kourimská, L.; Borkovcová, M.; Busina, T.; Adámek, M.; Bednárová, M.; Krajsa, J. Nutritional potential of selected insect species reared on the island of Sumatra. Int. J. Environ. Res. Public Health 2017, 14, 521. [Google Scholar] [CrossRef] [Green Version]
- Rodjaroen, S.; Thongprajukaew, K.; Khongmauang, P.; Malawa, S.; Tuntikawingwong, K.; Saekhow, S. Ontogenic development of digestive enzymes in mealworm larvae (Tenebrio molitor) and their suitable harvesting time for use as fish feed. Insects 2020, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Kulma, M.; Kouřimská, L.; Homolková, D.; Božik, M.; Plachý, V.; Vrabec, V. Effect of developmental stage on the nutritional value of edible insects. A case study with Blaberus craniifer and Zophobas morio. J. Food Compos. Anal. 2020, 92, 103570. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bordereau, C.; Andersen, S.O. Structural cuticular proteins in termite queens. Comp. Biochem. Physiol. B Comp. Biochem. 1978, 60, 251–256. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 29 February 2020).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: New York, NY, USA, 2016; pp. 1–260. [Google Scholar] [CrossRef]
- Patton, R.L. Growth and development parameters of Acheta domesticus. Ann. Entomol. Soc. Am. 1978, 7, 40–42. [Google Scholar] [CrossRef]
- McCluney, K.E.; Rishabh, C.D. The effects of hydration on growth of the house cricket, Acheta domesticus. J. Insect Sci. 2008, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, S.; Fratini, F.; Turchi, B.; Mattioli, S.; Bosco, A.D.; Tuccinardi, T.; Nozic, S.; Paci, G. Former foodstuff products in Tenebrio molitor rearing: Effects on growth, chemical composition, microbiological load, and antioxidant status. Animals 2019, 9, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Xie, B.; Dong, C.; Wang, M.; Liu, H. Can closed artificial ecosystem have an impact on insect microbial community? A case study of yellow mealworm (Tenebrio molitor L.). Ecolo. Engin. 2016, 86, 183–189. [Google Scholar] [CrossRef]
- Dossey, A.T.; Tatum, J.T.; McGill, W.L. Modern insect-based food industry: Current status, insect processing technology, and recommendations moving forward. In Insects as Sustainable Food Ingredients: Production, Processing and Food Applications, 1st ed.; Dossey, A.T., Morales-Ramos, M., Rojas, G., Eds.; Academic Press: London, UK, 2016; pp. 113–152. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, H.G.; Lee, K.Y.; Yoon, H.J.; Kim, N.J. Effects of agricultural byproducts, DDG and MSG, on the larval development of mealworms. Int. J. Ind. Entomol. 2016, 32, 69–79. [Google Scholar] [CrossRef]
- van Broekhoven, S.; Oonincx, D.G.; van Huis, A.; van Loon, J.J. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Collavo, A.; Glew, R.H.; Huang, Y.-S.; Chuang, L.-T.; Bosse, R.; Paoletti, M.G. House cricket small-scale farming. In Ecological Implications of Mini Livestock: Potential of Insects, Rodents, Frogs, and Snails; Paoletti, M.G., Ed.; Science Publishers, Inc.: Enfield, NH, USA, 2005; pp. 515–540. ISBN 1-57808-339-7. [Google Scholar]
- Vaga, M.; Berggren, A.; Pauly, T.; Jansson, A. Effect of red clover-only diets on house crickets (Acheta domesticus) growth and survival. J. Insects Food Feed 2020, 6, 179–189. [Google Scholar] [CrossRef]
- Quek, X.T.; Liang, L.; Tham, H.H.; Yeo, H.; Tan, M.K.; Tan, H.T.W. Are the growth and survival of Acheta domesticus comparable when reared on okara, waste vegetables and premium animal feed? J. Insects Food Feed 2019, 6, 161–168. [Google Scholar] [CrossRef]
- González, C.M.; Garzón, R.; Rosell, C.M. Insects as ingredients for bakery goods. A comparison study of H. illucens, A. domestica and T. molitor flours. Innov. Food Sci. Emerg. Technol. 2019, 51, 205–210. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.H.; Hong, S.-J.; Kim, N.J. Nutritional value of mealworm, Tenebrio molitor as food source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Busler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar] [CrossRef]
- Araujo, R.R.S.; dos Santos Benfica, T.A.R.; Ferraz, V.B.; Santos, E.M. Nutritional composition of insects Gryllus assimilis and Zophobas morio: Potential foods harvested in Brazil. J. Food Compos. Anal. 2019, 76, 22–26. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Veenenbos, M.E.; Oonincx, D.G.A.B. Carrot supplementation does not affect house cricket performance (Acheta domesticus). J. Insects Food Feed 2017, 3, 217–221. [Google Scholar] [CrossRef]
- Sorjonen, J.M.; Valtonen, A.; Hirvisalo, E.; Karhapää, M.; Lehtovaara, V.J.; Lindgren, J.; Marnila, P.; Mooney, P.; Mäki, M.; Siljander-Rasi, H.; et al. The plant-based by-product diets for the mass-rearing of Acheta domesticus and Gryllus bimaculatus. PLoS ONE 2019, 14, e0218830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachenicht, M.W.; Clusella-Trullas, S.; Boardman, L.; Le Roux, C.; Terblanche, J.S. Effects of acclimation temperature on thermal tolerance, locomotion performance and respiratory metabolism in Acheta domesticus L. (Orthoptera: Gryllidae). J. Insect Physiol. 2010, 56, 822–830. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | CF | CF/VW 1:9 | CF/GW 1:9 | CF/CM 1:9 | CF/HM 1:9 |
|---|---|---|---|---|---|
| Total organic carbon (C%) | 19.74 ± 3.38 b | 9.43 ± 1.71 d | 16.63 ± 2.25 c | 19.66 ± 1.70 b | 30.08 ± 2.16 a |
| Total nitrogen (N%) | 1.28 ± 0.12 a | 0.16 ± 0.01 d | 0.60 ± 0.05 b | 0.37 ± 0.03 c | 0.30 ± 0.03 c |
| Protein (g/kg) | 212.8 ± 8.31 a | 27.88 ± 4.78 e | 76.18 ± 7.74 b | 39.65 ± 7.11 d | 50.33 ± 5.84 e |
| Carbohydrate (g/kg) | 125.7 ± 1.74 a | 19.25 ± 0.70 b | 14.01 ± 1.03 d | 13.35 ± 0.71 d | 16.90 ± 0.93 c |
| Fat (g/kg) | 3.22 ± 1.08 c | 2.34 ± 0.99 c | 4.24 ± 0.70 b | 3.19 ± 0.64 c | 5.58 ± 0.60 a |
| Phosphorous (P%) | 0.43 ± 0.02 a | 0.10 ± 0.01 e | 0.18 ± 0.02 c | 0.16 ± 0.01 d | 0.21 ± 0.02 b |
| Potassium (K%) | 0.49 ± 0.04 b | 0.10 ± 0.05 d | 0.64 ± 0.06 a | 0.37 ± 0.04 c | 0.42 ± 0.05 c |
| Calcium (Ca%) | 1.02 ± 0.02 c | 0.97 ± 0.07 c | 1.38 ± 0.03 a | 1.15 ± 0.06 b | 0.79 ± 0.03 d |
| Components | First Dimension | Second Dimension | Third Dimension |
|---|---|---|---|
| Total organic carbon (C%) | 0.250 | 0.888 | −0.210 |
| Total nitrogen (N%) | 0.973 | −0.169 | 0.088 |
| Protein (g/kg) | 0.980 | −0.163 | −0.057 |
| Carbohydrate (g/kg) | 0.913 | −0.277 | −0.283 |
| Fat (g/kg) | 0.078 | 0.915 | 0.097 |
| Phosphorous (P%) | 0.982 | 0.045 | −0.154 |
| Potassium (K%) | 0.567 | 0.447 | 0.662 |
| Calcium (Ca%) | 0.052 | −0.278 | 0.948 |
| Components | CF | CF/VW 1:9 | CF/GW 1:9 | CF/CM 1:9 | CF/HM 1:9 |
|---|---|---|---|---|---|
| A. domesticus | |||||
| Crude protein (g/kg) | 67.25 ± 0.10 a | 61.20 ± 0.57 c | 65.30 ± 0.50 b | 57.80 ± 0.10 d | 56.40 ± 0.40 e |
| Crude fat (g/kg) | 14.41 ± 0.03 c | 17.10 ± 0.65 b | 19.30 ± 0.53 a | 18.60 ± 0.29 a | 19.40 ± 0.43 a |
| Fibre (g/kg) | 15.72 ± 0.03 c | 17.50 ± 0.66 b | 17.83 ± 0.38 b | 18.60 ± 0.29 a | 19.20 ± 0.37 a |
| Ash (g/kg) | 4.80 ± 0.04 b | 5.40 ± 0.54 b | 6.20 ± 0.39 a | 5.20 ± 0.32 b | 4.98 ± 0.32 b |
| Energy (MJ/kg) | 17.35 ± 0.10 e | 18.22 ± 0.01 d | 18.27 ± 0.02 c | 18.64 ± 0.01 b | 18.78 ± 0.02 a |
| T. molitor | |||||
| Crude protein (g/kg) | 47.18 ± 0.04 a | 46.30 ± 0.47 b | 42.30 ± 0.27 c | 38.92 ± 0.48 d | 37.90 ± 0.29 e |
| Crude fat (g/kg) | 43.08 ± 0.05 e | 43.30 ± 0.24 d | 45.20 ± 0.43 c | 46.70 ± 0.31 b | 47.50 ± 0.36 a |
| Fibre (g/kg) | 7.44 ± 0.02 d | 8.01 ± 0.37 c | 8.90 ± 0.41 b | 9.50 ± 0.23 ab | 9.30 ± 0.16 a |
| Ash (g/kg) | 3.08 ± 0.05 b | 3.01 ± 0.39 b | 3.20 ± 0.38 b | 4.85 ± 0.21 a | 5.30 ± 0.33 a |
| Energy (MJ/kg) | 24.17 ± 0.01 d | 24.39 ± 0.02 c | 24.59 ± 0.01 b | 24.68 ± 0.01 a | 24.60 ± 0.02 b |
| Z. morio | |||||
| Crude protein (g/kg) | 46.79 ± 1.03 a | 45.70 ± 0.48 b | 41.20 ± 0.31 c | 39.40 ± 0.34 d | 38.70 ± 0.39 d |
| Crude fat (g/kg) | 42.04 ± 0.74 d | 43.20 ± 0.31 c | 44.30 ± 0.37 b | 45.70 ± 0.41 a | 46.30 ± 0.42 a |
| Fibre (g/kg) | 9.26 ± 0.04 c | 9.43 ± 0.30 c | 11.30 ± 0.11 a | 10.20 ± 0.48 b | 9.32 ± 0.20 c |
| Ash (g/kg) | 2.61 ± 0.03 b | 2.89 ± 0.31 b | 3.01 ± 0.15 b | 4.70 ± 0.34 a | 4.89 ± 0.27 a |
| Energy (MJ/kg) | 24.10 ± 0.02 d | 24.43 ± 0.01 b | 24.38 ± 0.01 c | 24.75 ± 0.01 a | 24.75 ± 0.01 a |
| Components | First Dimension | Second Dimension |
|---|---|---|
| Crude protein (g/kg) | 0.929 | −0.304 |
| Crude fat (g/kg) | −0.985 | 0.160 |
| Fibre (g/kg) | 0.959 | 0.053 |
| Ash (g/kg) | 0.675 | 0.730 |
| Energy (MJ/100 g) | −0.992 | 0.105 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements. Insects 2020, 11, 604. https://doi.org/10.3390/insects11090604
Harsányi E, Juhász C, Kovács E, Huzsvai L, Pintér R, Fekete G, Varga ZI, Aleksza L, Gyuricza C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements. Insects. 2020; 11(9):604. https://doi.org/10.3390/insects11090604
Chicago/Turabian StyleHarsányi, Endre, Csaba Juhász, Elza Kovács, László Huzsvai, Richárd Pintér, György Fekete, Zsolt István Varga, László Aleksza, and Csaba Gyuricza. 2020. "Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements" Insects 11, no. 9: 604. https://doi.org/10.3390/insects11090604