Identification of Loci Controlling the Dwarfism Trait in the White Sailfin Molly (Poecilia latipinna) Using Genome-Wide Association Studies Based on Genotyping-By-Sequencing

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
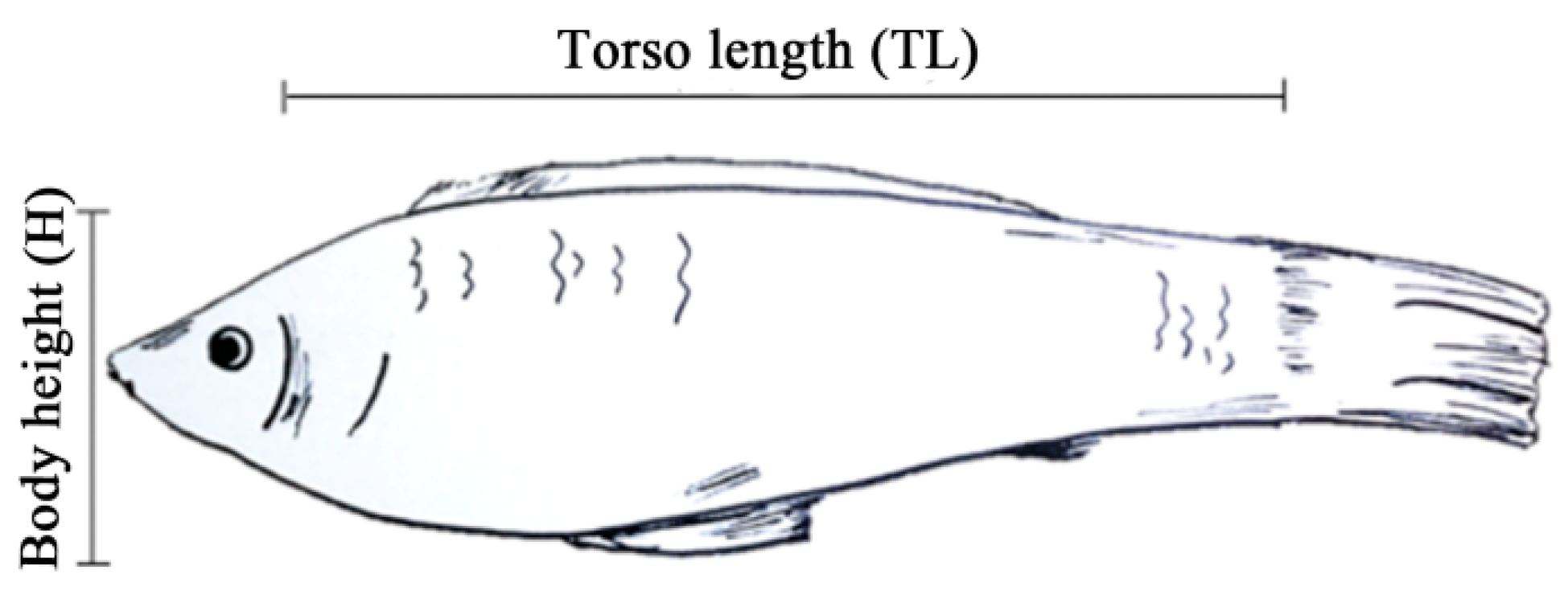
2.2. Animals and Phenotyping
2.3. Comparison of Skeletal Structures between the Dwarf Molly and the Normal Molly
2.4. Genotyping
2.5. Population Structural and Linkage Disequilibrium Analyses
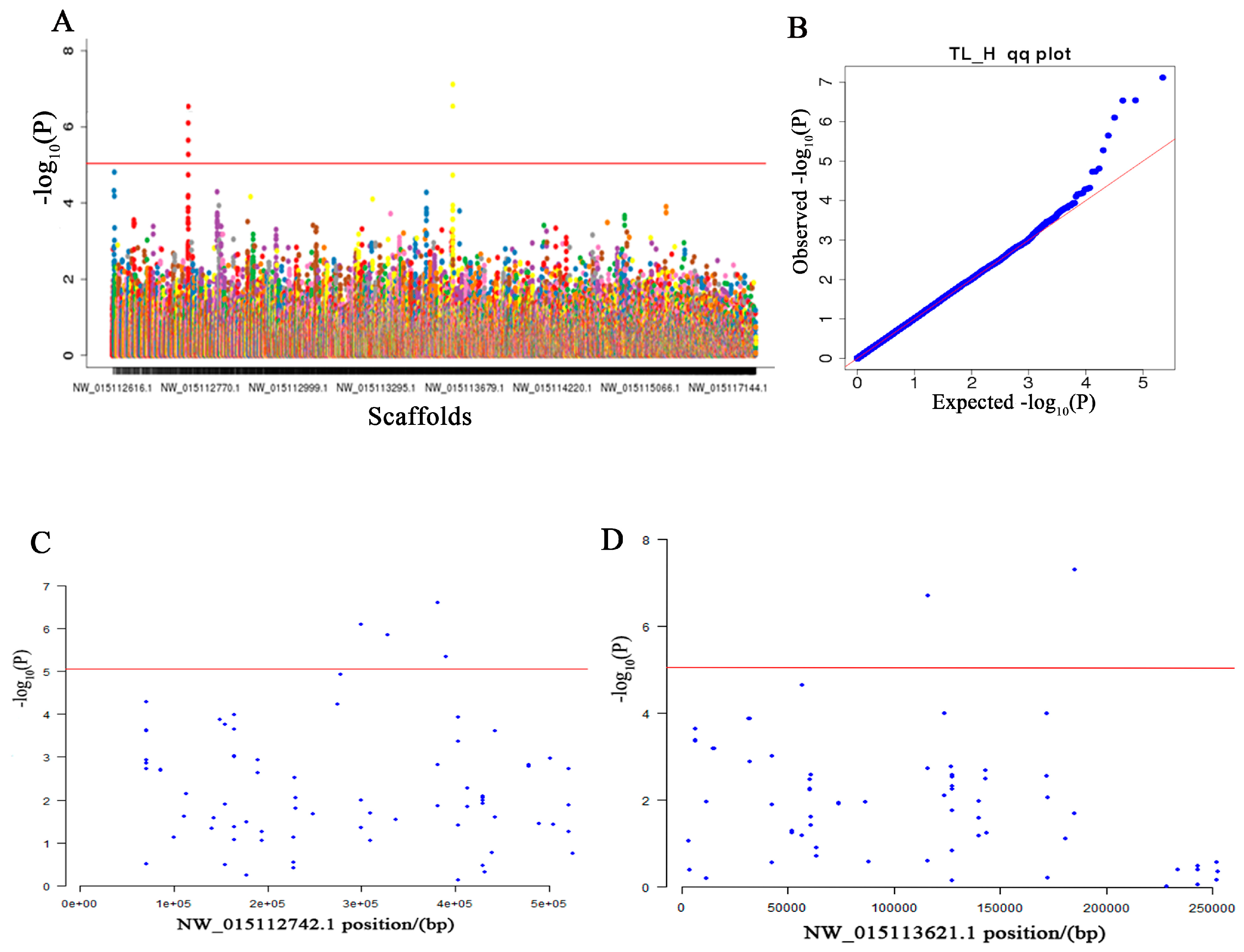
2.6. Genome-Wide Association Studies
2.7. RNA Extraction and Quantitative Real-Time PCR
3. Results and Discussion
3.1. Phenotypic Data of the White Molly
3.2. Comparison of Skeletal Structures between the Dwarf Molly and the Normal Molly
3.3. Sequencing Data Statistics and Quality Assessment
3.4. Single Nucleotide Polymorphism Filtering and Population Component Analysis
3.5. Genome-Wide Association Studies
3.6. Prediction of Candidate Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, K.B.; Liu, C.; Chang, O.Q.; Wang, F.; Ma, B.Y.; Wu, S.Q. The skeletal defects and inheritance in dwarf mutants of Xiphophorus helleri. J. Fish. China 2011, 35, 809–815. [Google Scholar] [CrossRef]
- Witten, P.E.; Gil-Martens, L.; Huysseune, A.; Takle, H.; Hjelde, K. Towards a classification and an understanding of developmental relationships of vertebral body malformations in Atlantic salmon (Salmo salar L.). Aquaculture 2009, 295, 6–14. [Google Scholar] [CrossRef]
- Haller, G.; McCall, K.; Jenkitkasemwong, S.; Sadler, B.; Antunes, L.; Nikolov, M.; Whittle, J.; Upshaw, Z.; Shin, J.; Baschal, E.; et al. A missense variant in SLC39A8 is associated with severe idiopathic scoliosis. Nat. Commun. 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Polak-Juszczak, L. Impact of strontium on skeletal deformities in Baltic cod (Gadus morhua callaris L.). Chemosphere 2011, 83, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Perrott, M.R.; Symonds, J.E.; Walker, S.P.; Preece, M.A.; Davie, P.S. Prevalence of spinal abnormalities in Chinook salmon smolt and influence of early rearing temperature and growth rates. J. Fish Dis. 2018, 41, 1111–1116. [Google Scholar] [CrossRef]
- Kupsco, A.; Schlenk, D. Molecular mechanisms of selenium-Induced spinal deformities in fish. Aquat. Toxicol. 2016, 179, 143–150. [Google Scholar] [CrossRef]
- Torres-Dowdall, J.; Karagic, N.; Plath, M.; Riesch, R. Evolution in caves: Selection from darkness causes spinal deformities in teleost fishes. Biol. Lett. 2018, 14, 20180197. [Google Scholar] [CrossRef]
- Chin, H.N.; Loh, R.; Hong, Y.C.; Gibson-Kueh, S. Case studies of spinal deformities in ornamental koi, Cyprinus carpio L. J. Fish Dis. 2017, 40, 65–71. [Google Scholar] [CrossRef]
- Tong, X.H.; Liu, Q.H.; Xu, S.H.; Ma, D.Y.; Xiao, Z.Z.; Xiao, Y.S.; Li, J. Skeletal development and abnormalities of the vertebral column and of the fins in hatchery-reared turbot Scophthalmus maximus. J. Fish. Biol. 2012, 80, 486–502. [Google Scholar] [CrossRef]
- Gedeon, A.K.; Tiller, G.E.; Le Merrer, M.; Heuertz, S.; Tranebjaerg, L.; Chitayat, D.; Robertson, S.; Glass, I.A.; Savarirayan, R.; Cole, W.G.; et al. The molecular basis of X-linked spondyloepiphyseal dysplasia tarda. Am. J. Hum. Genet. 2001, 68, 1386–1397. [Google Scholar] [CrossRef]
- Xia, X.Y.; Cui, Y.X.; Zhou, Y.C.; Zhou, X.; Shi, Y.C.; Wei, L.; Li, X.J.; Huang, Y.F.; Huang, T.T. A novel insertion mutation in the SEDL gene results in X-linked spondyloepiphyseal dysplasia tarda in a large Chinese pedigree. Clin. Chim. Acta 2009, 410, 39–42. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, S.K.; Minchin, J.E.N.; Gordon, T.N.; Rawls, J.F.; Parichy, D.M. Dwarfism and increased adiposity in the gh1 mutant Zebrafish vizzini. Endocrinology 2013, 154, 1476–1487. [Google Scholar] [CrossRef] [PubMed]
- Inohaya, K.; Takano, Y.; Kudo, A. Production of Wnt4b by floor plate cells is essential for the segmental patterning of the vertebral column in medaka. Development 2010, 137, 1807–1813. [Google Scholar] [CrossRef] [PubMed]
- Tenghe, A.M.M.; Bouwman, A.C.; Berglund, B.; Strandberg, E.; de Koning, D.J.; Veerkamp, R.F. Genome-wide association study for endocrine fertility traits using single nucleotide polymorphism arrays and sequence variants in dairy cattle. J. Dairy Sci. 2016, 99, 5470–5485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edea, Z.; Hong, J.K.; Jung, J.H.; Jung, J.H.; Kim, D.W.; Kim, Y.M.; Kim, E.S.; Shin, S.S.; Jung, Y.C.; Kim, K.S. Detecting selection signatures between Duroc and Duroc synthetic pig populations using high-density SNP chip. Anim. Genet. 2017, 48, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Siadjeu, C.; Mayland-Quellhorst, E.; Albach, D.C. Genetic diversity and population structure of trifoliate yam (Dioscorea dumetorum Kunth) in Cameroon revealed by genotyping-by-sequencing (GBS). BMC Plant Biol. 2018, 18, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Apodaca, J.J.; Trexler, J.C.; Jue, N.K.; Schrader, M.; Travis, J. Large-Scale Natural Disturbance Alters Genetic Population Structure of the Sailfin Molly, Poecilia latipinna. Am. Nat. 2013, 181, 254–263. [Google Scholar] [CrossRef]
- van Eeden, F.J.; Granato, M.; Schach, U.; Brand, M.; Furutani-Seiki, M.; Haffter, P.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; Kane, D.A.; et al. Mutations affecting somite formation and patterning in the zebrafish, Danio rerio. Development 1996, 123, 153–164. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Dong, S.S.; Xu, J.Y.; He, W.M.; Yang, T.L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of loci governing eight agronomic traits using a GBS-GWAS approach and validation by QTL mapping in soya bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xie, F.; Zhu, M.; Cao, Y.L.; Xiong, F. Correlation of SHOX gene deficiency and mutation with skeleton X-ray characteristics of idiopathic short stature patients. J. Third Mil. Med. Univ. 2011, 06, 604–607. [Google Scholar]
- Zhou, Z.H.; Shu, D.M.; Guo, K.Y.; Yang, C.F.; Xiong, Y. Study on the Inheritance Law of Yellow Feather Dwarf Chicken. Chin. J. Anim. Sci. 1998, 03, 26–27. [Google Scholar]
- Yun, S.F.; Yan, Z.L.; Li, C.P. The present situation of pathogenesis and genetics of short stature. Chin. J. Birth Health Hered. 2012, 03, 137–139. [Google Scholar]
- Afonso, J.M.; Montero, D.; Robaina, L.; Astorga, N.; Izquierdo, M.S.; Ginés, R. Association of a lordosis-scoliosis-kyphosis deformity in gilthead seabream (Sparus aurata) with family structure. Fish Physiol. Biochem. 2000, 22, 159–163. [Google Scholar] [CrossRef]
- Reich, D.; Price, A.L.; Patterson, N. Principal component analysis of genetic data. Nat. Genet. 2008, 40, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Su, I.I.; Tezuka, T.; Yasuda, T.; Mikoshiba, K.; Tarakhovsky, A.; Yamamoto, T. BANK regulates BCR-induced calcium mobilization by promoting tyrosine phosphorylation of IP(3) receptor. EMBO J. 2002, 21, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Wilbe, M.; Jokinen, P.; Truvé, K.; Seppala, E.H.; Karlsson, E.K.; Biagi, T.; Hughes, A.; Bannasch, D.; Andersson, G.; Hansson-Hamlin, H.; et al. Genome-wide association mapping identifies multiple loci for a canine SLE-related disease complex. Nat. Genet. 2010, 42, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Yang, W.; Zhao, M.; Mok, C.C.; Chan, T.M.; Wong, R.W.; Lee, K.W.; Mok, M.Y.; Wong, S.N.; Ng, I.O.; et al. Association of BANK1 and TNFSF4 with systemic lupus erythematosus in Hong Kong Chinese. Genes Immun. 2009, 10, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Hirohata, S.; Wang, L.W.; Miyagi, M.; Yan, L.; Seldin, M.F.; Keene, D.R.; Crabb, J.W.; Apte, S.S. Punctin, a novel ADAMTS-like molecule, ADAMTSL-1, in extracellular matrix. J. Biol. Chem. 2002, 277, 12182–12189. [Google Scholar] [CrossRef] [PubMed]
- Alazami, A.M.; Al-Owain, M.; Alzahrani, F.; Shuaib, T.; Al-Shamrani, H.; Al-Falki, Y.H.; Al-Qahtani, S.M.; Alsheddi, T.; Colak, D.; Alkuraya, F.S. Loss of function mutation in LARP7, chaperone of 7SK ncRNA, causes a syndrome of facial dysmorphism, intellectual disability, and primordial dwarfism. Hum. Mutat. 2012, 33, 1429–1434. [Google Scholar] [CrossRef]
- Hollink, I.H.; Alfadhel, M.; Al-Wakeel, A.S.; Ababneh, F.; Pfundt, R.; de Man, S.A.; Jamra, R.A.; Rolfs, A.; Bertoli-Avella, A.M.; van de Laar, I.M. Broadening the phenotypic spectrum of pathogenic LARP7 variants: Two cases with intellectual disability, variable growth retardation and distinct facial features. J. Hum. Genet. 2016, 61, 229–233. [Google Scholar] [CrossRef]
- Barboric, M.; Lenasi, T.; Chen, H.; Johansen, E.B.; Guo, S.; Peterlin, B.M. 7SK snRNP/P-TEFb couples transcription elongation with alternative splicing and is essential for vertebrate development. Proc. Natl. Acad. Sci. USA 2009, 106, 7798–7803. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | NWM | DWM |
|---|---|---|
| Torso Lenght (cm) | 3.6137 ± 0.0451 ** | 2.1828 ± 0.0451 |
| Height (cm) | 1.6264 ± 0.0211 * | 1.5666 ± 0.0211 |
| TL/H | 2.2256 ± 0.0244 ** | 1.4097 ± 0.0244 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, F.; Liu, J.; Ren, M.; Li, J.; Bao, H.; Wu, C. Identification of Loci Controlling the Dwarfism Trait in the White Sailfin Molly (Poecilia latipinna) Using Genome-Wide Association Studies Based on Genotyping-By-Sequencing. Genes 2019, 10, 418. https://doi.org/10.3390/genes10060418
Shao F, Liu J, Ren M, Li J, Bao H, Wu C. Identification of Loci Controlling the Dwarfism Trait in the White Sailfin Molly (Poecilia latipinna) Using Genome-Wide Association Studies Based on Genotyping-By-Sequencing. Genes. 2019; 10(6):418. https://doi.org/10.3390/genes10060418
Chicago/Turabian StyleShao, Fan, Jing Liu, Mengyuan Ren, Junying Li, Haigang Bao, and Changxin Wu. 2019. "Identification of Loci Controlling the Dwarfism Trait in the White Sailfin Molly (Poecilia latipinna) Using Genome-Wide Association Studies Based on Genotyping-By-Sequencing" Genes 10, no. 6: 418. https://doi.org/10.3390/genes10060418