Unveiling the Venom Composition of the Colombian Coral Snakes Micrurus helleri, M. medemi, and M. sangilensis

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
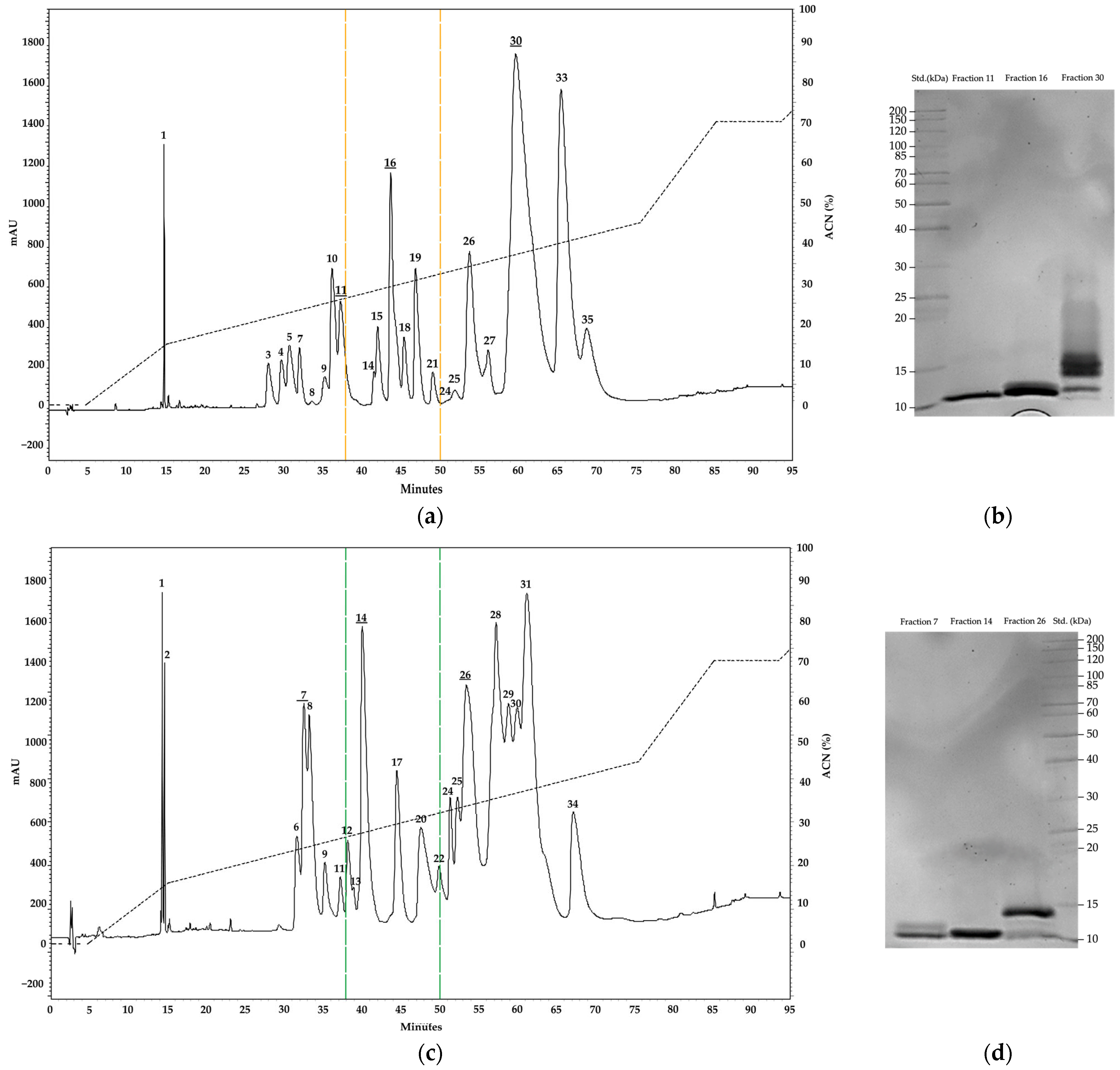
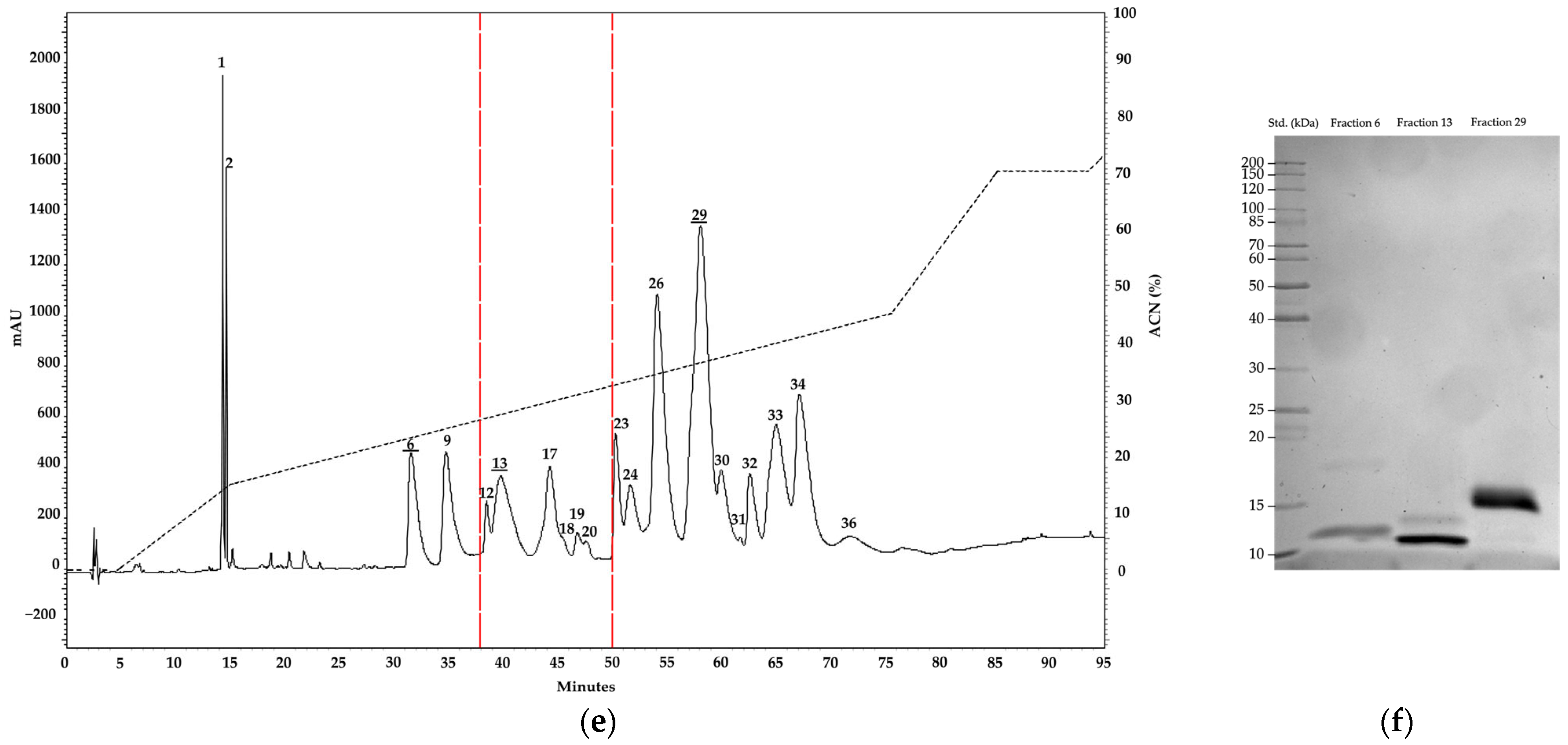
2.1. Electrophoresis and Chromatography Analysis
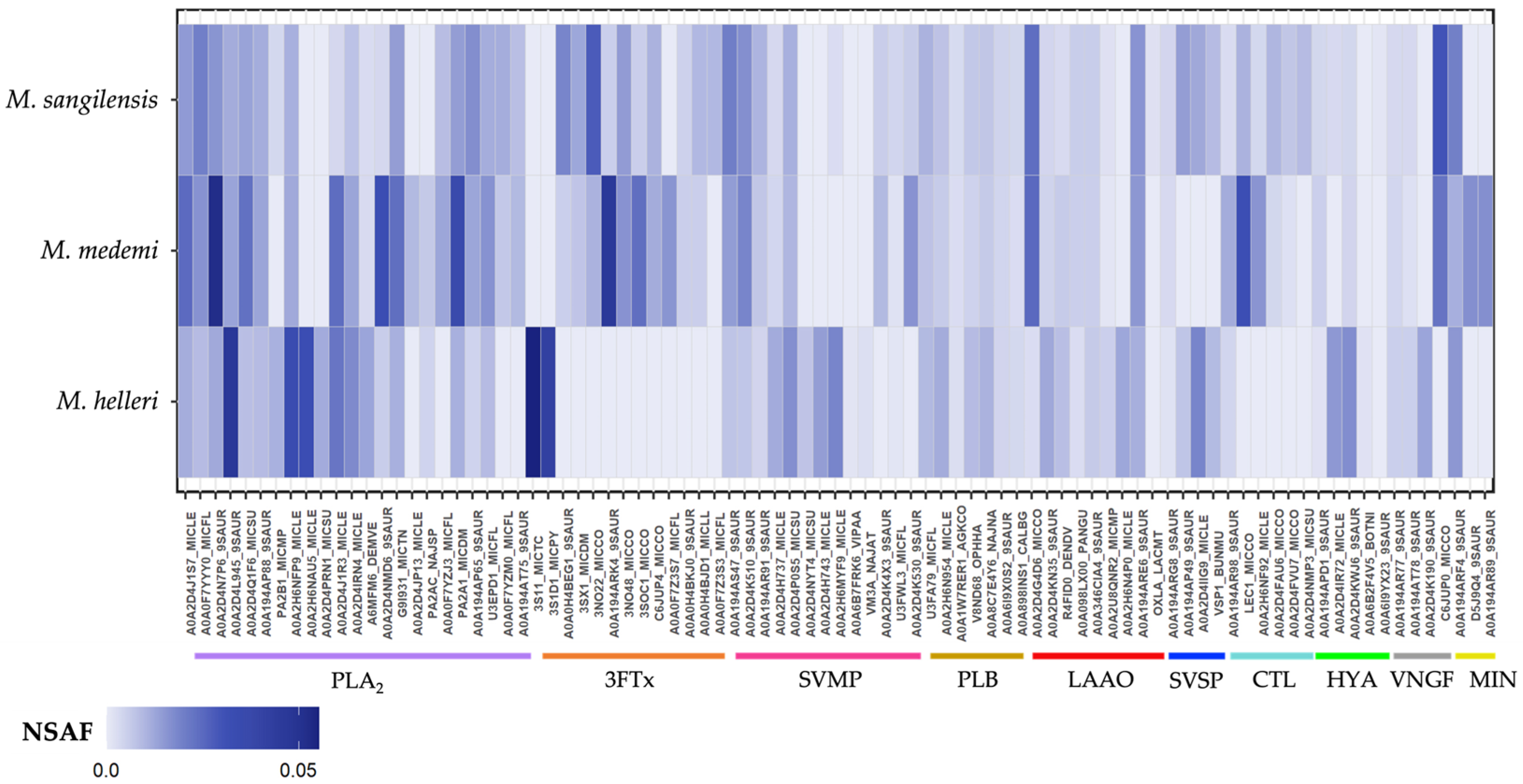
2.2. Protein Identification and Quantification
2.3. Enzymatic Activities
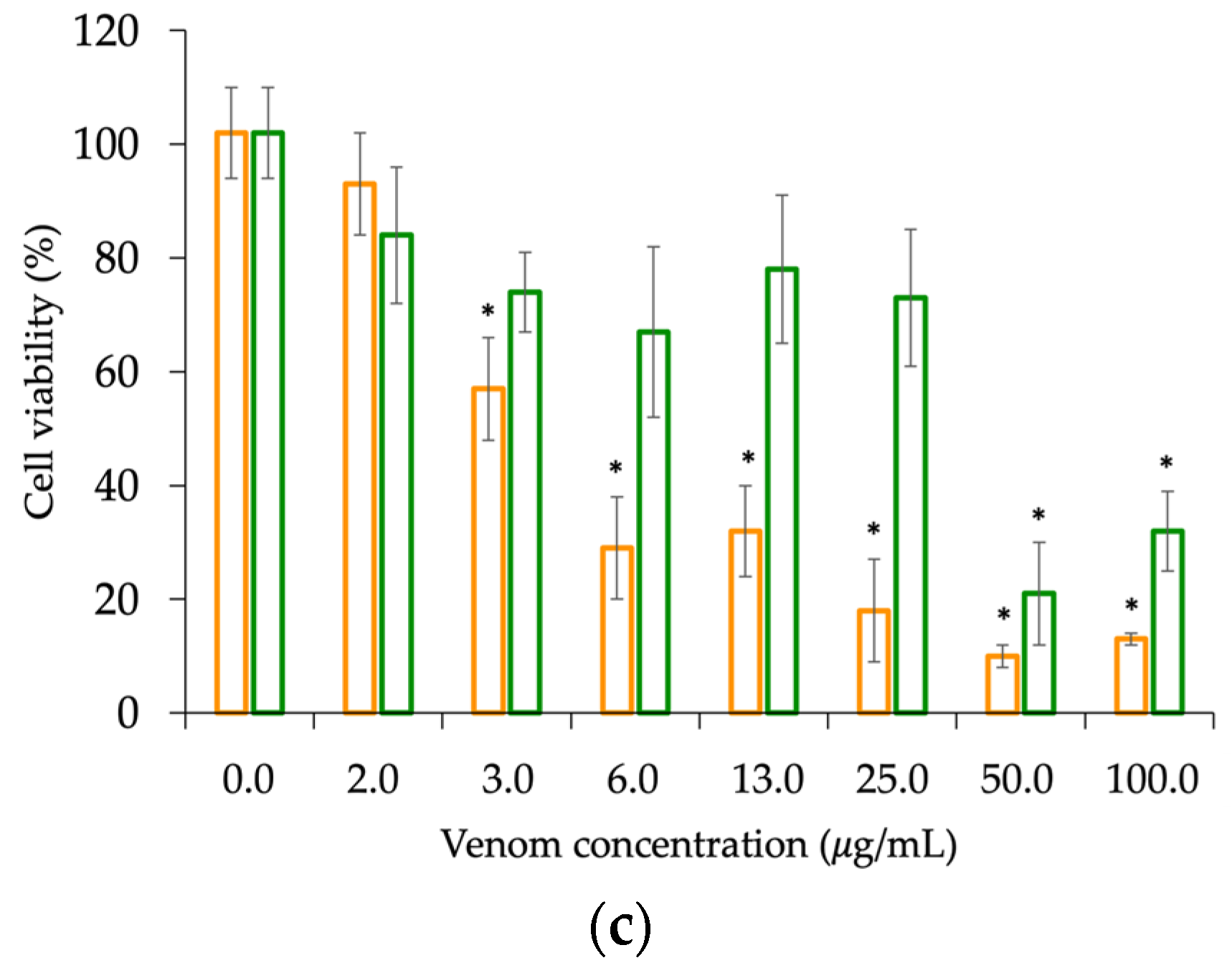
2.4. Venom Toxicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Target Species and Venoms
5.2. Experimental Animals
5.3. Venom Characterization
5.3.1. SDS-PAGE
5.3.2. Reversed-Phase Chromatography (RP-HPLC)
5.4. Protein Identification and Quantification
5.4.1. Protein Digestion with Trypsin
5.4.2. LC-MS/MS
5.4.3. Protein Identification
5.4.4. Protein Quantification
5.5. Biological Activities
5.5.1. Phospholipase A2 activity
5.5.2. Hyaluronidase Activity
5.5.3. Protease Activity
5.5.4. Lethal Dose
5.5.5. Cytotoxicity
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–2. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; De Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; De Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef]
- Chippaux, J.P. Incidence and mortality due to snakebite in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef]
- Gómez, J. Informe del Evento: Accidente Ofídico, Colombia. 2021. Available online: https://www.ins.gov.co/buscador-eventos/Informesdeevento/ACCIDENTEOFIDICOINFORME2021.pdf (accessed on 12 May 2023).
- Otero-Patiño, R. Snake Bites in Colombia BT-Clinical Toxinology in Australia, Europe, and Americas. In Clinical Toxinology in Australia, Europe, and Americas; Gopalakrishnakone, P., Vogel, C.W., Seifert, S.A., Tambourgi, D.V., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 3–50. ISBN 978-94-017-7438-3. [Google Scholar]
- Baldé, M.C.; Chippaux, J.-P.; Boiro, M.Y.; Stock, R.P.; Massougbodji, A. Use of antivenoms for the treatment of envenomation by Elapidae snakes in Guinea, Sub-Saharan Africa. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 6. [Google Scholar] [CrossRef] [PubMed]
- Hegde, R.P.; Rajagopalan, N.; Doley, R.; Kini, R.M. Snake Venom Three-Finger Toxins. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 287–302. ISBN 978-0-8493-9165-1. [Google Scholar]
- Ranawaka, U.; Lalloo, D.; de Silva, H. Neurotoxicity in Snakebite—The Limits of Our Knowledge. PLoS Negl. Trop. Dis. 2013, 7, e2302. [Google Scholar]
- Da Silva, N.J., Jr.; Bucaretchi, F. Mecanismo de açao do veneno elapídico e aspectos clínicos dos acidentes. In Animais Peçonhentos no Brasil; Cardoso, J., de Siqueira, F., Wen, F., Sant’Ana, C., Haddad, V., Eds.; Sarvier: São Paulo, Brazil, 2009; pp. 116–124. [Google Scholar]
- Gutiérrez, J.M. Snakebite Envenomation in Central America. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 491–507. [Google Scholar]
- Rey-Suárez, P.; Núñez, V.; Fernández, J.; Lomonte, B.; Nuñez, V.; Fernández, J.; Lomonte, B. Integrative characterization of the venom of the coral snake Micrurus dumerilii (Elapidae) from Colombia: Proteome, toxicity, and cross-neutralization by antivenom. J. Proteom. 2016, 136, 262–273. [Google Scholar] [CrossRef]
- Rey-Suárez, P.; Mónica, S.; Torres, U.; Marin-villa, M.; Lomonte, B.; Núñez, V. Novel three-finger toxins from Micrurus dumerilii and Micrurus mipartitus coral snake venoms: Phylogenetic relationships and characterization of Clarkitoxin-I-Mdum. Toxicon 2019, 170, 85–93. [Google Scholar]
- Arteaga, A.F.; Bustamante-Enríquez, L.; Guayasamin, J. The Amphibians and Reptiles of Mindo; Ecco Bella Beauty: Quito, Ecuador, 2013; ISBN 978-9942134967. [Google Scholar]
- Lynch, J. El contexto de las serpientes de Colombia con un análisis de las amenazas en contra de su conservación. Rev. la Acad. Colomb. Ciencias Exactas Físicas y Nat. 2012, 36, 435–449. [Google Scholar]
- Cadle, J.E.; Greene, H.W. Phylogenetic patterns, biogeography, and the ecological structure of Neotropical snake assemblages. In Species Diversity in Ecological Communities: Historical and Geographical Perspectives; Ricklefs, R.E., Schluter, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 281–293. [Google Scholar]
- Uetz, P.; Freed, P.; Aguilar, R.; Reyes, F.; Hošek, J. The Reptile Database. Available online: http://reptile-database.reptarium.cz (accessed on 12 May 2023).
- Roze, J.A. Coral Snakes of the Americas: Biology, Identification, and Venoms; Krieger Publishing Company: Malabar, FL, USA, 1996; ISBN 0-89464-847-0. [Google Scholar]
- Campbell, J.A.; Lamar, W. The Venomous Reptiles of the Western Hemisphere (Vol. 1); Comstock Publishing: New York, NY, USA, 2004; ISBN 978-0801441417. [Google Scholar]
- Caicedo-Portilla, J.R.; Lynch, J.D. Micrurus medemi. In Libro rojo de reptiles de Colombia (2015); Orales-Betancourt, M.A., Lasso, C.A., Páez, V.P., Bock, B.C., Eds.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Universidad de Antioquia: Bogotá, Colombia, 2015; pp. 98–100. ISBN 978-958-888-979-5. [Google Scholar]
- Montoya, A.; Díaz Flórez, R.A.; Mancera, D. A new departmental and elevational record for the Villavicencio Coralsnake, Micrurus medemi Roze 1967 (Squamata: Elapidae), in Colombia. Reptil. Amphib. 2022, 29, 189–190. [Google Scholar] [CrossRef]
- Angarita-Sierra, T.; Montaño-Londoño, L.F.; Bravo-Vega, C.A. ID please: Evaluating the utility of Facebook as a source of data for snake research and conservation. An. Acad. Bras. Cienc. 2022, 94, 1–25. [Google Scholar] [CrossRef]
- Niceforo-Maria, H. Los Ofidios de Colombia. Revista de La Academia Colombiana Ciencias Exactas Fisicas Y Naturales. Los Ofidios de Colombia 1942, 5, 89–101. [Google Scholar]
- Li, J.; Zhang, H.; Liu, J.; Xu, K. Novel genes encoding six kinds of three-finger toxins in Ophiophagus hannah (king cobra) and function characterization of two recombinant long-chain neurotoxins. Biochem. J. 2006, 398, 233–242. [Google Scholar] [CrossRef]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta 2006, 1761, 1246–1259. [Google Scholar] [CrossRef]
- Sartim, M.A.; Sampaio, S. V Snake venom galactoside-binding lectins: A structural and functional overview. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–11. [Google Scholar] [CrossRef]
- Serrano, S.M.T. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef]
- Olaoba, O.T.; Karina dos Santos, P.; Selistre-de-Araujo, H.S.; Ferreira de Souza, D.H. Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon X 2020, 7, 100052. [Google Scholar] [CrossRef]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.d.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W. A brief review of the scientific history of several lesser-known snake venom proteins: L-amino acid oxidases, hyaluronidases and phosphodiesterases. Toxicon 2013, 62, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Calvete, J. Strategies in “snake venomics” aiming at an integrative view of compositional, functional, and immunological characteristics of venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Pla, D.; Pérez, A.; Rodríguez, Y.; Zavaleta, A.; Salas, M.; Lomonte, B.; Calvete, J. Venomic analysis of the poorly studied desert coral snake, Micrurus tschudii tschudii, supports the 3FTx/PLA2 dichotomy across Micrurus venoms. Toxins 2016, 8, 178. [Google Scholar] [CrossRef]
- Olamendi-Portugal, T.; Batista, C.V.F.; Restano-Cassulini, R.; Pando, V.; Villa-Hernandez, O.; Zavaleta-Martínez-Vargas, A.; Salas-Arruz, M.C.; Rodríguez de la Vega, R.C.; Becerril, B.; Possani, L.D. Proteomic analysis of the venom from the fish eating coral snake Micrurus surinamensis: Novel toxins, their function and phylogeny. Proteomics 2008, 8, 1919–1932. [Google Scholar] [CrossRef]
- Rey-Suárez, P.; Núñez, V.; Gutiérrez, J.M.; Lomonte, B. Proteomic and biological characterization of the venom of the redtail coral snake, Micrurus mipartitus (Elapidae), from Colombia and Costa Rica. J. Proteom. 2011, 75, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Rey-Suárez, P.; Fernández, J.; Sasa, M.; Pla, D.; Vargas, N.; Bénard-Valle, M.; Sanz, L.; Corrêa-Netto, C.; Núñez, V.; et al. Venoms of Micrurus coral snakes: Evolutionary trends in compositional patterns emerging from proteomic analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Alape-Girón, A.; Angulo, Y.; Sanz, L.; Gutiérrez, J.M.; Calvete, J.; Lomonte, B.; Fernández, J.; Alape-Girón, A.; Angulo, Y.; et al. Venomic and Antivenomic Analyses of the Central American Coral Snake, Micrurus nigrocinctus (Elapidae). J. Proteome Res. 2011, 10, 1816–1827. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Quesada-Bernat, S.; Ramos, T.; Casais-e-Silva, L.L.; Corrêa-Netto, C.; Silva-Haad, J.J.; Sasa, M.; Lomonte, B.; Calvete, J.J. New insights into the phylogeographic distribution of the 3FTx/PLA 2 venom dichotomy across genus Micrurus in South America. J. Proteom. 2019, 200, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Altamirano, J.A.; Salazar-Valenzuela, D.; Medina-Villamizar, E.J.; Quirola, D.R.; Patel, K.; Vaiyapuri, S.; Lomonte, B.; Almeida, J.R. First Insights into the Venom Composition of Two Ecuadorian Coral Snakes. Int. J. Mol. Sci. 2022, 23, 14686. [Google Scholar] [CrossRef]
- Mena, G.; Chaves-Araya, S.; Chacón, J.; Török, E.; Török, F.; Bonilla, F.; Sasa, M.; Gutiérrez, J.M.; Lomonte, B.; Fernández, J. Proteomic and toxicological analysis of the venom of Micrurus yatesi and its neutralization by an antivenom. Toxicon X 2022, 13, 100097. [Google Scholar] [CrossRef]
- Olamendi-Portugal, T.; Batista, C.V.F.; Pedraza-Escalona, M.; Restano-Cassulini, R.; Zamudio, F.Z.; Benard-Valle, M.; Rafael de Roodt, A.; Possani, L.D. New insights into the proteomic characterization of the coral snake Micrurus pyrrhocryptus venom. Toxicon 2018, 153, 23–31. [Google Scholar] [CrossRef]
- Lomonte, B.; Calvete, J.; Davinia, P.; Rey-Suárez, P.; Sanz, L.; Gutiérrez, J.; Sasa, M. Venomic analyses of coralsnakes. In The Biology of the Coralsnakes; da Silva, N., Porras, L., Aird, S., Prudente, A., Eds.; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2021; pp. 485–518. ISBN 978-0-9720154-6-2. [Google Scholar]
- Castillo-Beltrán, M.C.; Hurtado-Gómez, J.P.; Corredor-Espinel, V.; Ruiz-Gómez, F.J. A polyvalent coral snake antivenom with broad neutralization capacity. PLoS Negl. Trop. Dis. 2018, 13, e0007250. [Google Scholar] [CrossRef]
- Davies, E.L.; Arbuckle, K. Coevolution of snake venom toxic activities and diet: Evidence that ecological generalism favours toxicological diversity. Toxins 2019, 11, 711. [Google Scholar] [CrossRef]
- Tanaka, G.D.; Furtado, M.D.F.D.; Portaro, F.C.V.; Sant’Anna, O.A.; Tambourgi, D.V. Diversity of Micrurus snake species related to their venom toxic effects and the prospective of antivenom neutralization. PLoS Negl. Trop. Dis. 2010, 4, e622. [Google Scholar] [CrossRef]
- Da Silva, N.J.; Aird, S.D. Prey Specificity, Comparative Lethality and Compositional Differences of Coral Snake Venoms. Comp. Biochem. Physiol. Part C 2001, 128, 425–456. [Google Scholar]
- Calvete, J.J. Snake venomics at the crossroads between ecological and clinical toxinology. Biochemist 2019, 41, 28–33. [Google Scholar] [CrossRef]
- Casais-e-Silva, L.L.; Teixeira, C.F.P.; Lebrun, I.; Lomonte, B.; Alape-Girón, A.; Gutiérrez, J.M. Lemnitoxin, the major component of Micrurus lemniscatus coral snake venom, is a myotoxic and pro-inflammatory phospholipase A2. Toxicol. Lett. 2016, 257, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Lippa, E.; Török, F.; Gómez, A.; Corrales, G.; Chacón, D.; Sasa, M.; Gutiérrez, J.M.; Lomonte, B.; Fernández, J. First look into the venom of Roatan Island’s critically endangered coral snake Micrurus ruatanus: Proteomic characterization, toxicity, immunorecognition and neutralization by an antivenom. J. Proteom. 2019, 198, 177–185. [Google Scholar] [CrossRef]
- Rey-Suárez, P.; Acosta, C.; Torres, U.; Saldarriaga-Córdoba, M.; Lomonte, B.; Núñez, V. MipLAAO, a new L-amino acid oxidase from the redtail coral snake Micrurus mipartitus. PeerJ 2018, 6, e4924. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B. Caracterización biológica y proteómica del veneno de Micrurus mipartitus en Colombia y aislamiento de la toxina mayoritaria. Actual. Biológicas 2012, 34, 145–146. [Google Scholar]
- Bénard-Valle, M.; Carbajal-Saucedo, A.; de Roodt, A.; López-Vera, E.; Alagón, A. Biochemical characterization of the venom of the coral snake Micrurus tener and comparative biological activities in the mouse and a reptile model. Toxicon 2014, 77, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Rey-Suárez, P.; Floriano, R.S.; Rostelato-Ferreira, S.; Saldarriaga-Córdoba, M.; Núñez, V.; Rodrigues-Simioni, L.; Lomonte, B. Mipartoxin-I, a novel three-finger toxin, is the major neurotoxic component in the venom of the redtail coral snake Micrurus mipartitus (Elapidae). Toxicon 2012, 60, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Rey-Suárez, P.; Núñez, V.; Saldarriaga-Córdoba, M.; Lomonte, B. Primary structures and partial toxicological characterization of two phospholipases A 2 from Micrurus mipartitus and Micrurus dumerilii coral snake venoms. Biochimie 2017, 137, 88–98. [Google Scholar] [CrossRef]
- Cecchini, A.L.; Marcussi, S.; Silveira, L.B.; Borja-Oliveira, C.R.; Rodrigues-Simioni, L.; Amara, S.; Stábeli, R.G.; Giglio, J.R.; Arantes, E.C.; Soares, A.M. Biological and enzymatic activities of Micrurus sp. (Coral) snake venoms. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 140, 125–134. [Google Scholar] [CrossRef]
- Terra, A.L.C.; Moreira-Dill, L.S.; Simões-Silva, R.; Monteiro, J.R.N.; Cavalcante, W.L.G.; Gallacci, M.; Barros, N.B.; Nicolete, R.; Teles, C.B.G.; Medeiros, P.S.M.; et al. Biological characterization of the Amazon coral Micrurus spixii snake venom: Isolation of a new neurotoxic phospholipase A2. Toxicon 2015, in press. [CrossRef]
- Bucaretchi, F.; De Capitani, E.M.; Vieira, R.J.; Rodrigues, C.K.; Zannin, M.; Da Silva, N.J.; Casais-e-Silva, L.L.; Hyslop, S. Coral snake bites (Micrurus spp.) in Brazil: A review of literature reports. Clin. Toxicol. 2016, 54, 222–234. [Google Scholar] [CrossRef]
- Guo, C.; Liu, S.; Yao, Y.; Zhang, Q.; Sun, M.Z. Past decade study of snake venom l-amino acid oxidase. Toxicon 2012, 60, 302–311. [Google Scholar] [CrossRef]
- Sanz, L.; de Freitas-Lima, L.N.; Quesada-Bernat, S.; Graça-de-Souza, V.K.; Soares, A.M.; Calderón, L.d.A.; Calvete, J.J.; Caldeira, C.A.S. Comparative venomics of Brazilian coral snakes: Micrurus frontalis, Micrurus spixii spixii, and Micrurus surinamensis. Toxicon 2019, 166, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Gómez, J.P.; Vargas-Ramírez, M.; Ruíz Gómez, F.J.; Fouquet, A.; Fritz, U. Multilocus phylogeny clarifies relationships and diversity within the Micrurus lemniscatus. Salamandra 2021, 57, 229–239. [Google Scholar]
- Da Silva, N.; Griffin, P.R.; Aird, S.D. Comparative chromatography of Brazilian coral snake (Micrurus venoms. Comp. Biochem. Physiol. Part B Biochem. 1991, 100, 117–126. [Google Scholar] [CrossRef]
- Pereañez, J.A.; Núñez, V.; Patiño, A.C.; Mónica, L.; Quintana, J.C. Inhibitory effects of plant phenolic compounds on enzymatic and cytotoxic activities induced by a snake venom phospholipase A2. Vitae 2011, 18, 295–304. [Google Scholar] [CrossRef]
- Fernández, J.; Vargas-Vargas, N.; Pla, D.; Sasa, M.; Rey-Suárez, P.; Sanz, L.; Gutiérrez, J.M.; Calvete, J.; Lomonte, B. Snake venomics of Micrurus alleni and Micrurus mosquitensis from the Caribbean region of Costa Rica reveals two divergent compositional patterns in New World elapids. Toxicon 2015, 107, 217–233. [Google Scholar] [CrossRef]
- Mackessy, S. Venom Composition in Rattlesnakes: Trends and Biological Significance. In The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 495–510. [Google Scholar]
- Dashevsky, D.; Bénard-Valle, M.; Neri-Castro, E.; Youngman, N.J.; Zdenek, C.N.; Alagón, A.; Portes-Junior, J.A.; Frank, N.; Fry, B.G. Anticoagulant Micrurus venoms: Targets and neutralization. Toxicol. Lett. 2021, 337, 91–97. [Google Scholar] [CrossRef]
- Sant’Ana Malaque, C.; Gutiérrez, J.M. Snakebite Envenomation in Central and South America. In Critical Care Toxicology; Brent, J., Burkhart, K., Dargan, P., Hatten, B., Megarbane, B., Palmer, R., Eds.; Springer: Cham, Switzerland, 2015; pp. 1–22. ISBN 9783319207902. [Google Scholar]
- Manock, S.R.; Suarez, G.; Graham, D.; Avila-Aguero, M.L.; Warrell, D.A. Neurotoxic envenoming by South American coral snake (Micrurus lemniscatus helleri): Case report from eastern Ecuador and review. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1127–1132. [Google Scholar] [CrossRef]
- Da Silva, A.M.; da Fonseca, W.L.; de Araujo Valente Neto, E.; Bisneto, P.F.; Contreras-Bernal, J.; Sachett, J.; Monteiro, W.M.; Bernarde, P.S. Envenomation by Micrurus annellatus bolivianus (Peters, 1871) coral snake in the western Brazilian Amazon. Toxicon 2019, 166, 34–38. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M.T. Structural considerations of the snake venom metalloproteinases, key members of the M12 reprolysin family of metalloproteinases. Toxicon 2005, 45, 969–985. [Google Scholar] [CrossRef]
- Kini, R.M. Serine proteases affecting blood coagulation and fibrinolysis from snake venoms. Pathophysiol. Haemost. Thromb. 2006, 34, 200–204. [Google Scholar] [CrossRef]
- Renjifo, C.; Smith, E.N.; Hodgson, W.C.; Renjifo, J.M.; Sanchez, A.; Acosta, R.; Maldonado, J.H.; Riveros, A. Neuromuscular activity of the venoms of the Colombian coral snakes Micrurus dissoleucus and Micrurus mipartitus: An evolutionary perspective. Toxicon 2012, 59, 132–142. [Google Scholar] [CrossRef]
- Ramos, H.R.; Vassão, R.C.; de Roodt, A.R.; Santos E Silva, E.C.; Mirtschin, P.; Ho, P.L.; Spencer, P.J. Cross neutralization of coral snake venoms by commercial Australian snake antivenoms. Clin. Toxicol. 2017, 55, 33–39. [Google Scholar] [CrossRef]
- Oliveira, D.A.; Harasawa, C.; Seibert, C.S.; Casais e Silva, L.L.; Pimenta, D.C.; Lebrun, I.; Sandoval, M.R.L. Phospholipases A2 isolated from Micrurus lemniscatus coral snake venom: Behavioral, electroencephalographic, and neuropathological aspects. Brain Res. Bull. 2008, 75, 629–639. [Google Scholar] [CrossRef]
- Thangam, R.; Gunasekaran, P.; Kaveri, K.; Sridevi, G.; Sundarraj, S.; Paulpandi, M.; Kannan, S. A novel disintegrin protein from Naja naja venom induces cytotoxicity and apoptosis in human cancer cell lines in vitro. Process Biochem. 2012, 47, 1243–1249. [Google Scholar] [CrossRef]
- Utkin, Y.N. Three-finger toxins, a deadly weapon of elapid venom—Milestones of discovery. Toxicon 2013, 62, 50–55. [Google Scholar] [CrossRef]
- Shanbhag, V.K.L. Applications of snake venoms in treatment of cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 275–276. [Google Scholar] [CrossRef]
- De Carvalho, N.D.; Garcia, R.C.; Kleber Ferreira, A.; Rodrigo Batista, D.; Carlos Cassola, A.; Maria, D.; Lebrun, I.; Mendes Carneiro, S.; Castro Afeche, S.; Marcourakis, T.; et al. Neurotoxicity of coral snake phospholipases A2 in cultured rat hippocampal neurons. Brain Res. 2014, 1552, 1–16. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Sarkar, A. Cytotoxic Effects of Snake Venoms. In Snake Venoms, Toxinology; Gopalakrishnakone, P., Inagaki, H., Mukherjee, A.K., Rahmy, T.R., Vogel, C.-W., Eds.; Springer: Dordrecht, Germany, 2017; pp. 1–7. [Google Scholar]
- World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Inmunoglubolines; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- World Health Organization. WHO Annex 5. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins Replacement of Annex 2 of WHO Technical Report Series, No. 964; WHO: Geneva, Switzerland, 2017; pp. 197–388. [Google Scholar]
- Roze, J.A. A Check List of the New World Venomous Coral Snakes (Elapideae), with Descriptions of New Forms; American Museum of Natural History: New York, NY, USA, 1967; pp. 1–59. [Google Scholar]
- Smith, P.; Krohn, R.; Hermanson, G.; Mallia, A.; Gartner, F.; Provenzano, M.; Fujimoto, E.; Goeke, N.; Olson, B.; Klenk, D. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Lamberts, B. Use of coomassie brilliant blue R250 for the electrophoresis of microgram quantities of parotid saliva proteins on acrylamide-gel strips. Biochim. Biophys. Acta 1965, 107, 144–145. [Google Scholar] [CrossRef]
- Switzer, R.C., III; Merril, C.R.; Shifrin, S. A Highly Sensitive Silver Stain for Detecting Proteins and Peptides in Polyacrylamide Gels. Anal. Biochem. 1979, 98, 231–237. [Google Scholar] [CrossRef]
- Henao Duque, A.M.; Núñez Rangel, V. Maintenance of red-tail coral snake (Micrurus mipartitus) in captivity and evaluation of individual venom variability. Acta Biológica Colomb. 2016, 21, 593. [Google Scholar] [CrossRef]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Zybailov, B.; Mosley, A.L.; Sardiu, M.E.; Coleman, M.K.; Florens, L.; Washburn, M.P. Statistical analysis of membrane proteome expression changes in Saccharomyces cerevisiae. J. Proteome Res. 2006, 5, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- De Araújo, A.L.; Radvanyi, F. Determination of phospholipase A2 activity by a colorimetric assay using a pH indicator. Toxicon 1987, 25, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Memar, B.; Jamili, S.; Shahbazzadeh, D.; Bagheri, P.K. The first report on coagulation and phospholipase A2 activities of Persian Gulf lionfish, Pterois russelli, an Iranian venomous fish. Toxicon 2016, 113, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, M.A.; Navarro-Duque, C.; Varela-Julia, M.; Alagon, A.C. Molecular mass determination and assay of venom hyaluronidases by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Toxicon 1992, 30, 925–930. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef]
- Spearman, C. The method of ‘right and wrong cases’ (‘constant stimuli’) without Gauss’s formulae. Br. J. Psychol. 1904–1920 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn. Schmiedebergs. Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Guisande-Gonzáles, C.; Vaamonde Liste, A.; Barreiro-Felpeto, A. RWizard 2014, 1–452. Available online: http://www.ipez.es/RWizard/ (accessed on 26 May 2023).
- Corrêa-Netto, C.; Junqueira-De-Azevedo, I.d.L.M.; Silva, D.A.; Ho, P.L.; Leitão-De-Araújo, M.; Alves, M.L.M.; Sanz, L.; Foguel, D.; Zingali, R.B.; Calvete, J. Snake venomics and venom gland transcriptomic analysis of Brazilian coral snakes, Micrurus altirostris and M. corallinus. J. Proteom. 2011, 74, 1795–1809. [Google Scholar] [CrossRef]
- Lomonte, B.; Sasa, M.; Rey-Suárez, P.; Bryan, W.; Gutiérrez, J.M.J.M. Venom of the coral snake Micrurus clarki: Proteomic profile, toxicity, immunological cross-neutralization, and characterization of a three-finger Toxin. Toxins 2016, 8, 138. [Google Scholar] [CrossRef]
- Vergara, I.; Pedraza-Escalona, M.; Paniagua, D.; Restano-Cassulini, R.; Zamudio, F.; Batista, C.V.F.; Possani, L.D.; Alagón, A. Eastern coral snake Micrurus fulvius venom toxicity in mice is mainly determined by neurotoxic phospholipases A2. J. Proteom. 2014, 105, 295–306. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Retention Time | Relative Abundance | ||
|---|---|---|---|---|
| M. helleri | M. medemi | M. sangilensis | ||
| 1 | 14.3 | 0.9 | 2.7 | 3.8 |
| 2 | 14.6 | 0.0 | 1.0 | 1.5 |
| 3 | 28.0 | 1.5 | 0.0 | 0.0 |
| 4 | 29.7 | 1.4 | 0.0 | 0.0 |
| 5 | 30.7 | 2.5 | 0.0 | 0.0 |
| 6 | 31.6 | 0.0 | 2.8 | 8.1 |
| 7 | 32.3 | 1.4 | 3.7 | 0.0 |
| 8 | 33.4 | 0.1 | 2.7 | 0.0 |
| 9 | 35.1 | 1.2 | 2.3 | 7.3 |
| 10 | 36.2 | 4.3 | 0.0 | 0.0 |
| 11 | 37.3 | 4.8 | 1.0 | 0.0 |
| 12 | 38.3 | 0.0 | 1.8 | 1.0 |
| 13 | 39.3 | 0.0 | 0.2 | 5.7 |
| 14 | 40.8 | 0.4 | 15.9 | 0.0 |
| 15 | 42.0 | 0.2 | 0.0 | 0.0 |
| 16 | 43.7 | 7.9 | 0.0 | 0.0 |
| 17 | 44.3 | 0.0 | 5.8 | 3.4 |
| 18 | 45.4 | 1.9 | 0.0 | 0.2 |
| 19 | 46.8 | 4.4 | 0.0 | 0.7 |
| 20 | 47.5 | 0.0 | 7.2 | 0.1 |
| 21 | 49.0 | 0.8 | 0.0 | 0.0 |
| 22 | 49.8 | 0.0 | 0.9 | 0.0 |
| 23 | 50.2 | 0.0 | 0.0 | 3.3 |
| 24 | 51.5 | 0.0 | 2.2 | 2.3 |
| 25 | 52.1 | 0.4 | 1.4 | 0.0 |
| 26 | 53.7 | 7.0 | 12.6 | 14.9 |
| 27 | 56.1 | 2.0 | 0.0 | 0.0 |
| 28 | 57.2 | 0.0 | 12.2 | 0.0 |
| 29 | 58.4 | 0.0 | 1.2 | 23.3 |
| 30 | 59.8 | 35.9 | 0.9 | 1.9 |
| 31 | 61.2 | 0.0 | 11.6 | 0.0 |
| 32 | 62.6 | 0.0 | 0.0 | 3.6 |
| 33 | 65.2 | 17.1 | 0.0 | 9.3 |
| 34 | 67.1 | 0.0 | 9.9 | 8.6 |
| 35 | 68.8 | 3.7 | 0.0 | 0.0 |
| 36 | 71.8 | 0.0 | 0.0 | 1.1 |
| Species | LD50 (μg/Mice) | CI95% | Source |
|---|---|---|---|
| M. dumerilli | 23.72 | 18.10–31.10 | [41] |
| M. helleri (sensu lato) | 22.87 | 17.79–29.40 | |
| M. isozonus | 6.29 | 5.14–7.69 | |
| M. mipartitus | 33.62 | 26.10–43.20 | |
| M. spixii | 13.89 | Not determined | |
| M. surinamensis | 29.17 | 24.42–34.84 | |
| M. helleri | 22.87 | 17.79–29.4 | |
| M. medemi | 8.79 | 7.16–10.80 | |
| M. sangilensis | 15.85 | 11.50–21.70 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Vargas, A.; Franco-Vásquez, A.M.; Bolívar-Barbosa, J.A.; Vega, N.; Reyes-Montaño, E.; Arreguín-Espinosa, R.; Carbajal-Saucedo, A.; Angarita-Sierra, T.; Ruiz-Gómez, F. Unveiling the Venom Composition of the Colombian Coral Snakes Micrurus helleri, M. medemi, and M. sangilensis. Toxins 2023, 15, 622. https://doi.org/10.3390/toxins15110622
Rodríguez-Vargas A, Franco-Vásquez AM, Bolívar-Barbosa JA, Vega N, Reyes-Montaño E, Arreguín-Espinosa R, Carbajal-Saucedo A, Angarita-Sierra T, Ruiz-Gómez F. Unveiling the Venom Composition of the Colombian Coral Snakes Micrurus helleri, M. medemi, and M. sangilensis. Toxins. 2023; 15(11):622. https://doi.org/10.3390/toxins15110622
Chicago/Turabian StyleRodríguez-Vargas, Ariadna, Adrián Marcelo Franco-Vásquez, Janeth Alejandra Bolívar-Barbosa, Nohora Vega, Edgar Reyes-Montaño, Roberto Arreguín-Espinosa, Alejandro Carbajal-Saucedo, Teddy Angarita-Sierra, and Francisco Ruiz-Gómez. 2023. "Unveiling the Venom Composition of the Colombian Coral Snakes Micrurus helleri, M. medemi, and M. sangilensis" Toxins 15, no. 11: 622. https://doi.org/10.3390/toxins15110622