
Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions


1
Centre d’Ecologie Fonctionelle et Evolutive CEFE, Université Montpellier, CNRS, EPHE, IRD, 34293 Montpellier, France
2
Grupo de Ecoloxía Animal (GEA), Universidade de Vigo, 36310 Vigo, Spain
3
Department de Biodiversidad, Ecología y Evolución, Facultad de Ciencias Biologicas, Universidad Complutense de Madrid, José Antonio Novais, 2, 28040 Madrid, Spain
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(1), 30; https://doi.org/10.3390/d14010030
Submission received: 9 December 2021
/
Revised: 23 December 2021
/
Accepted: 24 December 2021
/
Published: 4 January 2022
(This article belongs to the Special Issue Molecular Phylogeny and Evolution of Earthworms)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Earthworm systematics have been limited by the small number of taxonomically informative morphological characters and high levels of homoplasy in this group. However, molecular phylogenetic techniques have yielded significant improvements in earthworm taxonomy in the last 15 years. Several different approaches based on the use of different molecular markers, sequencing techniques, and compromises between specimen/taxon coverage and phylogenetic information have recently emerged (DNA barcoding, multigene phylogenetics, mitochondrial genome analysis, transcriptome analysis, targeted enrichment methods, and reduced representation techniques), providing solutions to different evolutionary questions regarding European earthworms. Molecular phylogenetics have led to significant advances being made in Lumbricidae systematics, such as the redefinition or discovery of new genera (Galiciandrilus, Compostelandrilus, Vindoboscolex, Castellodrilus), delimitation and revision of previously existing genera (Kritodrilus, Eophila, Zophoscolex, Bimastos), and changes to the status of subspecific taxa (such as the Allolobophora chaetophora complex). These approaches have enabled the identification of problems that can be resolved by molecular phylogenetics, including the revision of Aporrectodea, Allolobophora, Helodrilus, and Dendrobaena, as well as the examination of small taxa such as Perelia, Eumenescolex, and Iberoscolex. Similar advances have been made with the family Hormogastridae, in which integrative systematics have contributed to the description of several new species, including the delimitation of (formerly) cryptic species. At the family level, integrative systematics have provided a new genus system that better reflects the diversity and biogeography of these earthworms, and phylogenetic comparative methods provide insight into earthworm macroevolution. Despite these achievements, further research should be performed on the Tyrrhenian cryptic complexes, which are of special eco-evolutionary interest. These examples highlight the potential value of applying molecular phylogenetic techniques to other earthworm families, which are very diverse and occupy different terrestrial habitats across the world. The systematic implementation of such approaches should be encouraged among the different expert groups worldwide, with emphasis on collaboration and cooperation.
1. Introduction
Earthworm systematics have been riddled since their inception by the limited number of morphological characters offered by the soft-bodied and conserved body plan of these organisms, with many of them being symplesiomorphic or homoplasious [1]. This has been reflected by an unstable taxonomy, even at higher taxonomic levels. For instance, at least seven different genus systems have been proposed for Lumbricidae throughout the 20th century [2,3,4,5,6,7,8] and the family system for Megascolecoidea is still subject to changes [9].
The advent of molecular phylogenetic techniques provided researchers with a new tool to obtain revised systematics, which would reflect the evolutionary relationships of earthworms (the aim of modern cladistics).
2. Molecular Phylogenetics Techniques
One of the first breakthroughs in earthworm molecular phylogenetics was the implementation of DNA barcoding (cytochrome C oxidase subunit I sequencing, COI) [10]. COI was introduced as a candidate for a universal molecular marker by [11], and it was subsequently accepted in this role for most animal taxa due to the robustness of its universal primers [12] and great range of phylogenetic signal, which allows the discrimination of closely related species and genetic lineages within single species. The use of this tool led to the discovery of the widespread presence of species-level cryptic lineages across the Lumbricidae [13,14], a pattern found to be the norm within different earthworm families [15,16]. COI barcodes have a high potential for phylogeographic analyses, in which a wide representation of individuals from different populations are required to infer genetic diversity and dispersion patterns (Figure 1). Application of the technique to whole earthworm communities has enabled comparative phylogeographic studies [17], in an approach which could help to decipher the different dispersal and colonization patterns of earthworms and their potential relationships with functional traits, geological history, and anthropogenic influence.
Although COI barcoding is a useful tool, it has been found to be limited for resolving phylogenetic relationships at above species level [18]. Other mitochondrial (such as COII, 16S, 12S, ND1) and nuclear molecular markers (such as 28S or 18S) showed similar limitations when used individually [19,20]. Their combination in multigene or multilocus phylogenetic analyses (Figure 1) has improved the ability of phylogenetic analysis to recover closely related species within monophyletic clades [21,22,23], even revealing some relationships at above genus (or even family) levels. Nonetheless, in some cases of ancient divergence or rapid radiation, this approach has been shown to be lacking power.
As a source of several molecular markers with the added value of establishing gene order, mitochondrial genome analysis represents an interesting alternative to multigene phylogenetic analysis [24,25]. Besides its advantages, such as increased above species level resolution due to increased number of markers and rare rearrangements, mitochondrial genomes overlook nuclear information, which could constitute a problem in cases of incomplete lineage sorting or heteroplasmy [26,27].
The obvious solution to the limitations presented by these methods would lie in the representation of the whole genome of a species within the phylogenetic analyses. As whole-genome sequencing (WGS) is unaffordable in several phylogenetic applications (particularly when a wide taxon coverage is required), transcriptomics appears to be the best alternative. Two studies have demonstrated the potential application of transcriptomics to earthworm molecular phylogenetics [28,29]; phylogenetic resolution at the family level and above was significantly increased, but at the cost of generating very complex bioinformatic pipelines for selecting the most informative orthologous genes. In addition, genes evolving at different rates can potentially support different topologies [28]. Another possible disadvantage of this methodology is the requirement of freshly preserved specimens, as well as the need for special preservation protocols and careful handling of RNA (relative to DNA) during the process.
Some of the most recent additions to the molecular phylogenetics toolbox include targeted enrichment methods, namely ultra-conserved elements (UCE [30]) and anchored hybrid enrichment (AHE [31]) (Figure 1). These techniques rely on previously existing genomes as the starting point for loci selection and probe design, chosen based on conservation and uniqueness through a sliding window approach [31]; this enables the capture of a consistent marker data set for all the taxa studied, while avoiding phylogenetically misleading parts of the genome (e.g., paralogs and pseudogenes) [32]. These techniques are an efficient, inexpensive way of sequencing hundreds (e.g., 609 in [27]) of orthologs, which in the case of AHE provide phylogenetic signal at both deep and shallow scale analyses [31]. They have displayed the ability to resolve problematic nodes in other animals [33,34]. The latter technique has been successfully applied to clitellates [35] and earthworms [27] and is currently being implemented in the Lumbricidae (Rafinesque-Schmaltz, 1815) by D.F. Marchán and collaborators.
All the aforementioned phylogenomic approaches are suitable for reconstructing phylogenetic relationships above the species level, but owing to the cost and the level of genetic variability they capture, they are not suitable for phylogeographic studies, in which several representatives per locality are required. Reduced representation techniques such as RAD-seq [36] and GBS [37] have been developed for this purpose (Figure 1). Both approaches share the same methodological basis, in which restriction enzymes are used to cut the genomic DNA, and the resulting fragments are sorted by size and sequenced. After bioinformatic treatment, these yield thousands of single nucleotide polymorphisms (SNPs), which are suitable for population genetics, selection signature analysis and cryptic lineage delimitation. For some examples of these approaches in earthworms, see [38,39,40,41].
Time-calibrated phylogenies can provide valuable information about divergence time and to test hypothesis about historical biogeography and environment-trait evolution correlations. Currently, Bayesian methods, such as the one implemented in BEAST [42], allows a wide range of substitution models (which can be different and independent for different sets of sites), flexible model specification, and choice of priors on parameters. Thus, these methods allow to estimate the time-calibrated trees, which are better adjusted to the molecular data analyzed and the calibrations provided. In the case of earthworms, this is the weak point of divergence time estimation; as soft bodied invertebrates, no direct body fossils exist for them, which can be used for calibration. Trace fossils (such as cocoons and galleries) and closely related annelid fossils have been implemented as a compromise [43], but the vast temporal scale and deep genetic divergence between the outgroups and the ingroup resulted in wide confidence intervals, which require cautious interpretation of results. Paleogeographic events and their correlation with splits between sister taxa have been implemented as an alternative to fossils [44,45], yet this approach has been criticized as relying on the assumption of vicariance; divergence between taxa being older than the paleogeographic event cannot be ruled out, resulting in divergence time estimation that are in practice only a minimum estimate. External substitution rates (obtained from previous analyses) should be used with caution as substitution rates for the same gene change between taxa even within the same family [22]; they often come from vicariance-based analyses [46] but they have also been obtained from more robust fossil-based analyses [47,48].
Besides the limitation of calibration sources, the choice of molecular technique can improve divergence time estimation; genomes, transcriptomes, and targeted enrichment methods provide hundreds or thousands of loci among which it is possible to choose the most suitable ones (clock-like or tree-like [49]).
3. Contributions of Molecular Phylogenetics to Lumbricidae Systematics
3.1. Advances
The multigene phylogenetic analysis conducted by [50] provided a clearer picture of the state of Lumbricidae systematics and highlighted some of the most obvious problems. For example, several lumbricid genera were recovered as paraphyletic or polyphyletic (particularly Aporrectodea (Orley 1885), Allolobophora (Eisen 1873), Dendrobaena (Eisen 1873), and Helodrilus (Hoffmeister 1845)), while others appeared to be monophyletic (for instance Eisenia (Malm 1877), Scherotheca (Bouché 1972), Bimastos (Moore 1893), Lumbricus (Linnaeus 1758), and Prosellodrilus (Bouché 1972)).
Within this framework, other rare or endemic genera which were absent or underrepresented in the aforementioned study have gradually been added in recent molecular phylogenetic studies (Figure 2). For example, [23] described and redefined two new genera from the northwestern Iberian Peninsula (Galiciandrilus (Dominguez et al. 2017) and Compostelandrilus (Dominguez et al. 2017)), which appeared as closely related taxa to the Balearic Postandrilus (Qiu and Bouche 1998). Interestingly, backward displaced male pores (from the usual position in segment 15) were observed in all representatives of Postandrilus, but not in all representatives of Galiciandrilus and Compostelandrilus. This finding, together with the fact that Cernosvitovia (Omodeo 1956) (with backward displaced pores) is phylogenetically unrelated, confirmed the suspicions of this character being homoplasious and thus devoid of systematic value.
These three genera have been recently found to be related to Kritodrilus (Bouche 1972) (whose type species was included in molecular analyses for the first time (Marchán et al. 2021a)) and surprisingly, to the newly described Central European genus Vindoboscolex [48]. The unlikely phylogenetic relationship of those isolated genera has provided some new insight into the early evolution of the Lumbricidae; an early branching clade would have occupied a wide area ranging from Galicia to Hungary and, subsequently, been fragmented into relict, geographically restricted clades.
The important inclusion of generotypes (type species of a genus) in molecular phylogenetic analyses provided other significant advances (Figure 2), such as restricting the genus Eophila (Rosa 1893) to the closest relatives of Eophila tellinii (Rosa 1888) (Eophila gestroi (Cognetti de Martiis 1905) and Eophila crodabepis (Paoletti et al. 2016)), which ended the story of the genus as a taxonomic wastebasket [47]. Similarly, the inclusion of Zophoscolex atlanticus (Bouché 1969) together with several other species previously attributed to Zophoscolex (Qiu and Bouche, 1998) restricted this genus to the French species, while the Iberian species were found to constitute the separate genus Castellodrilus [51].
Ref. [50] demonstrated the close relationship between the poorly known French species Allolobophora chaetophora (Bouché 1972) and Helodrilus cortezi (Qiu and Bouché 1998), which formed a well-supported clade separated from other representatives of their (previously considered) congeneric species. Avelona ligra (Bouché 1969) was shown as a sister species of All. chaetophora by [29] but with a very restricted number of Lumbricidae taxa included in the analysis. Ref. [52] included representatives of most of the previously described subspecies of All. chaetophora, He. musicus (Qiu and Bouché 1998) and Av. ligra in a multigene phylogenetic analysis, showing that H. cortezi, H. musicus, and All. chaetophora constitute a genus-level clade, Gatesona, which also includes the former subspecies of All. chaetophora now elevated to the species level (Figure 2). At the same time Gatesona was retrieved as the sister clade to Avelona, which together constitute a French lineage with most of its diversity restricted to the Massif Central.
Based on wide taxonomic sampling and multigene phylogenetic analysis, [53] recovered the cosmopolitan genera Dendrodrilus (Omodeo 1956) and Allolobophoridella (Mrsic 1990) within the same clade as the North American endemic species of Bimastos, indicating the synonymy of the former with the latter and their ancestral origin in North America together with their sister taxa Eisenoides (Gates 1969). In addition, the morphologically similar Healyella (Omodeo and Rota 1989) and Spermophorodrilus (Bouché 1975) were found to be phylogenetically unrelated and nested within a Dendrobaena sensu lato clade.
3.2. Remaining Questions
All the advances stressed both the suitability of molecular phylogenetic approaches and the need for their application to remaining systematic questions within the family.
One of the most glaring systematic issues is the taxonomic wastebaskets identified by [50] as non-monophyletic.
The first of these is Aporrectodea, which includes several of the most widespread and common lumbricids, such as Aporrectodea caliginosa (Savigny 1826), Aporrectodea trapezoides (Duges 1828), and Aporrectodea rosea (Savigny 1826). Furthermore, Ap. trapezoides (the generotype), Ap. caliginosa, and other species included by Bouché (1972) within Nicodrilus (Bouché 1972) appeared to form a well-supported lineage [21] clearly unrelated to Ap. rosea, Ap. georgii (Michaelsen 1890), or Ap. jassyensis (Michaelsen 1891) (Figure 2), all of which had been assigned to the genus Koinodrilus (Qiu and Bouché 1998). However, the latter species behave as rogue taxa in phylogenetic analyses; this means that their positions in phylogenetic trees are unstable and cannot be resolved with certainty. The addition of further representatives of Koinodrilus could allow the phylogenetic relationships of these taxa to stabilize; alternatively, the application of other molecular phylogenetic techniques with greater resolution at deeper nodes, such as AHE, could provide a solution to this problem. As all the Aporrectodea species (including, but not restricted to, Koinodrilus) not related to Ap. trapezoides cannot be considered to belong to a monophyletic Aporrectodea, they must be placed within newly defined genera once their phylogenetic relationships are finally resolved.
Allolobophora represents a similar case. The type species, Allolobophora chlorotica (Savigny 1826), appears to form a monophyletic clade with other green-pigmented lumbricids such as Allolobophora dubiosa (Orley 1881) and Allolobophora molleri (Rosa 1889) [50]. This clade appears to be unrelated to several Carpatho-Balkanic species (such as Allolobophora mehadiensis (Rosa 1895), Allolobophora robusta (Rosa 1895), and Allolobophora sturanyi dacica (Pop 1938)), which have previously been assigned to different genera (such as Serbiona and Karpatodinariona). Interestingly, the latter species appeared to be more closely related to Cernosvitovia [50] (Figure 2). The inclusion of more representatives of those Carpatho-Balkanic clades will be necessary to confirm the hypothesis that these “Allolobophora” forms belong to a redefined Cernosvitovia.
Some representatives of both Aporrectodea and Allolobophora present an additional challenge; deep species-level lineages or cryptic species that have been identified within Ap. trapezoides and Ap. caliginosa [21,54], Ap.rosea [55] and All. chlorotica [13,56]. In these cases, although the molecular phylogenetic evidence already exists, the taxonomy is lagging. The difficulty in describing morphologically cryptic lineages as species may explain why such work has not already been performed (but see [40,57] for alternative approaches to this task), in addition to the difficulty of including holotypes (or topotypes) of previously described taxa in order to assign valid names to these genetic lineages. Although daunting, this task must be undertaken; the aforementioned species are amongst the most abundant in anthropogenic habitats, such as crops and orchards, and all agroecological, ecotoxicological, and applied research targeting them should be accurately assigned to the relevant species, not to a loosely related complex of genetic lineages.
The same problem affects the genera Helodrilus and Dendrobaena. Some representatives of both have been included in molecular phylogenetic analyses [50,53,58,59] revealing that they consist of several unrelated genus-level clades (Figure 2). Although generotypes have been included in both cases, further representatives of their wide taxonomic diversity in Eastern Europe remain to be studied, and the molecular markers must be standardized in order to combine the fragmentary data. In the case of Dendrobaena, a name has already been proposed for species more closely related to Dendrobaena byblica (Rosa 1893) than to Dendrobaena octaedra (Savigny 1826): Omodeoia (Kvavadze 1994). Hence, if other species assigned to Omodeoia were recovered together with D. byblica in a monophyletic clade, there would be molecular support for division of the Dendrobaena species complex into Dendrobaena and Omodeoia. Interestingly, the newly described genus Phylomontanus (Bozorgui et al. 2019) appears to be closely related to D. byblica (Figure 2); a more comprehensive molecular phylogenetics study would enable testing whether this constitutes a junior synonym of Omodeoia or if it is a third lineage of “Dendrobaena-like” earthworms. In the case of Helodrilus, no replacement name appears to be available for species not closely related to Helodrilus oculatus (Hoffmeister 1895) (the type species). Thus, new genus names will need to be proposed for the other unrelated clades.
In addition to the most conspicuous genera, other smaller ones still pose important challenges for molecular phylogenetics. For example, Perelia (Easton 1983) includes several species of the Middle East and Central Asia, with a few Eastern Europe representatives. The phylogenetic relationships of these species between each other and with other lumbricid genera are currently unknown; in addition, one species was proposed to belong to the new genus Rhiphaeodrilus (Csuzdi and Pavlicek 2005), based on nephridia morphology. Incorporation of these species in a molecular phylogenetic framework would not only be helpful for the systematic revision of the Lumbricidae, but also for historical biogeographic reconstructions, as very few endemic representatives from beyond Eastern Europe are currently available. Eumenescolex (Qiu and Bouché 1998) is a poorly known genus of Western Mediterranean earthworms, with a strongly disjunct distribution (France, Italy, Spain), which is otherwise consistent with the geological history of the region. Members of the genus have been suggested to be related to Scherotheca [60], but they differ strikingly in body size and lifestyle from most species, other than the Corsican endemics. Molecular phylogenetics data on these elusive earthworms could provide more information on the intriguing role of Corsica, Sardinia, and Southern France in the evolution of lumbricids before they drifted apart in the Oligocene [61]. Scherotheca itself is a diverse genus, for which relatively little molecular phylogenetics data is available (but see [60] for four additional representatives, including a newly described one). In this case, molecular phylogenetics would serve as a starting point for determining their ancestral range (Spain, mainland France, or Corsica) and morphological radiation towards a giant anecic phenotype. The case of Eiseniona (Omodeo 1956) or Iberoscolex (Qiu and Bouché 1998) it is also worthy of mention that several Iberian species were described as belonging to the Eastern European genus Eiseniona, (which has also been considered synonym of Aporrectodea), but they were assigned by Qiu and Bouché (1998) to the genus Iberoscolex. The addition of the generotypes Iberoscolex microepigeus (Qiu and Bouché 1998) and Eiseniona handlirschi (Rosa 1897) to the representatives previously included in molecular phylogenetics analyses (Domínguez et al. 2015) will enable a statement to be made regarding the validity of these controversial genera [62].
4. Contributions of Molecular Phylogenetics to the Hormogastridae Systematics
4.1. Advances
The application of molecular phylogenetics to a comparatively small but well represented family in the Western Mediterranean, Hormogastridae (Michaelsen 1900) led to a drastic change in their systematics.
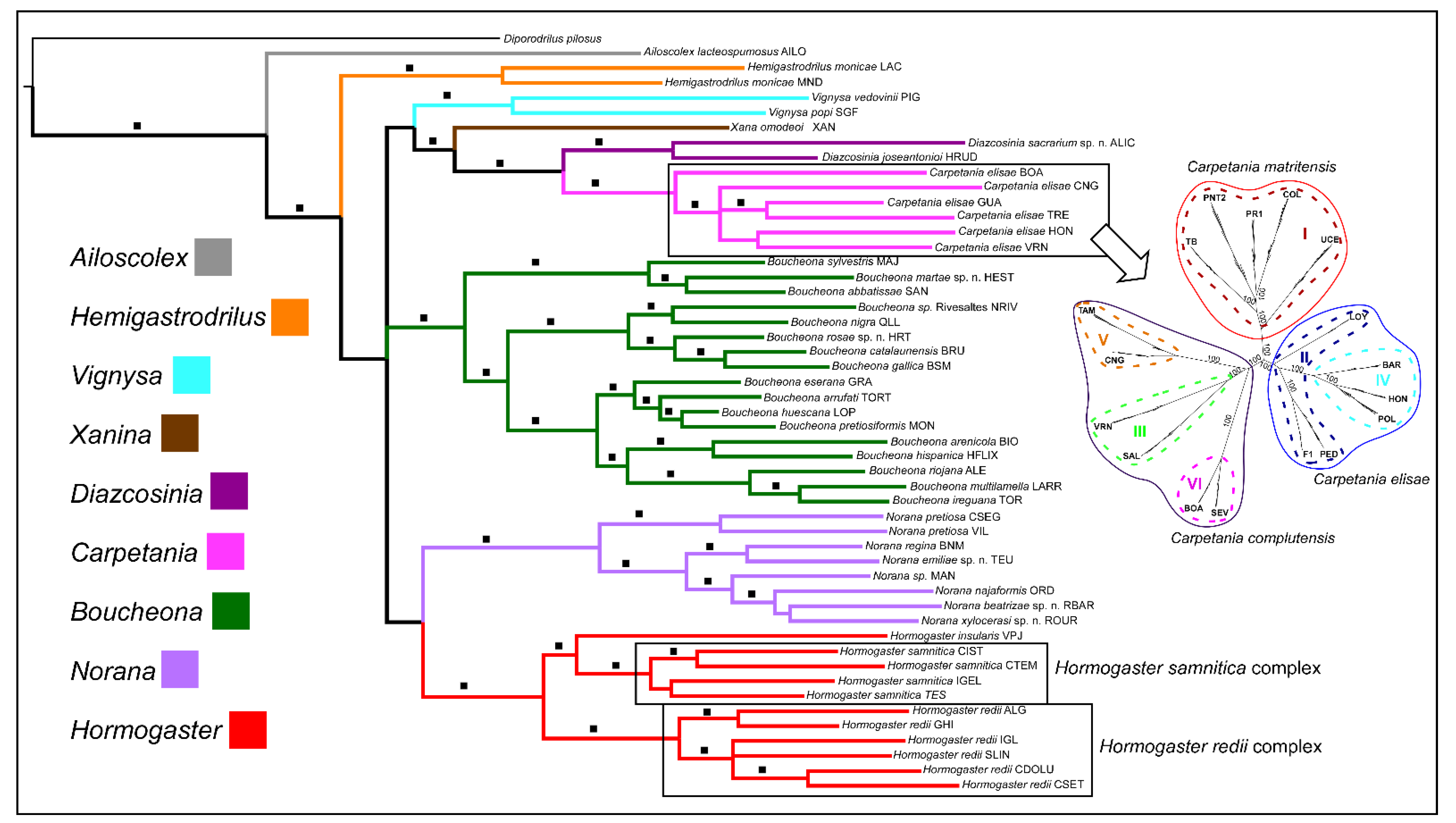
Integrative systematics (combining molecular phylogenetics and new morphological characters) led to the description of nine new species in six years [63,64,65,66], which constitutes an increase of 43%. Furthermore, molecular phylogenetic approaches uncovered high levels of cryptic speciation within the Hormogastridae [15]; the geographically restricted Hormogaster elisae (Álvarez 1977) complex (recently redescribed as the genus Carpetania Marchán et al. 2018) became an ideal model for studying the phylogeography of cryptic lineages [43] and the evolutionary processes involved in their diversification [39], culminating in the integrative description of the three component species [40] (Figure 3).
The reconstruction of an explicit phylogenetic framework through multigene methods enabled the application of the phylogenetic comparative method (rarely used in invertebrates) to study macroevolutionary patterns in Hormogastridae; in this way, the origin and radiation of a key evolutionary innovation (the multilamellar typhlosole) was linked to increasing body weight, with soil characteristics as secondary evolutionary pressures [67].
The systematics of the family were revealed to be in dire need of revision by Novo et al. [22,44], as the genus Hormogaster (Rosa 1887) appeared to include at least four independent, genus-level clades (later described as Hormogaster, Boucheona (Marchán et al. 2018), Norana (Marchán et al. 2018) and Carpetania (Marchán et al. 2018)), and different species-level taxa were described under the same name (such as Hormogaster praetiosa (Michaelsen 1899)). Transcriptomics analysis led to the inclusion of the highly divergent genera Ailoscolex (Bouché 1969) (previously thought to belong to a different family) and Hemigastrodrilus (Bouché 1970) as the earliest branching clades of the family [28]. However, it was the integration of morphological characters (including new ones [68]) and molecular markers that finally led to proposal of a revised genus system for Hormogastridae [69] (Figure 3). The new systematics of this family revealed a more complex outline of the diversity and biogeography of its genera, opening the door to future evolutionary and ecological research
Figure 3.
Modified from [66,69]. Bayesian inference of the phylogenetic relationships of Hormogastridae based on a combined morphological and molecular matrix (concatenated sequence of COI-16S-tRNAs-28S-H3 markers). The inset shows the species delimitation of the Carpetania elisae complex based on a maximum likelihood analysis of the SNP dataset obtained with GBS (modified from [39,40]).
Figure 3.
Modified from [66,69]. Bayesian inference of the phylogenetic relationships of Hormogastridae based on a combined morphological and molecular matrix (concatenated sequence of COI-16S-tRNAs-28S-H3 markers). The inset shows the species delimitation of the Carpetania elisae complex based on a maximum likelihood analysis of the SNP dataset obtained with GBS (modified from [39,40]).

4.2. Remaining Questions
Ref. [70] revealed two cryptic lineage complexes within the Tyrrhenian Hormogaster redii (Rosa 1887) and Hormogaster samnitica (Cognetti de Martiis, 1914); in a similar way to the former Hormogaster elisae complex, these species-level genetic lineages could not be easily delimited using multigene phylogenetic inference (Figure 3). The application of reduced representation techniques such as GBS and RAD-seq, together with increased sampling effort across their ranges, would probably enable integrative species delimitation and description within those complexes. This is not a minor issue; with some of the widest ranges among Hormogastridae and occupying a diverse range of habitats, H. redii and H. samnitica appear to possess adaptive and colonizing potential far beyond their hormogastrid kin. More precise knowledge of their systematics will enable these species to be used as evolutionary models to apply molecular evolution approaches, such as those implemented by [66,71].
5. Remaining Questions of Lumbricoidea
As outlined above, extensive advances have been made in knowledge and understanding of the main families of Lumbricoidea (Lumbricidae and Hormogastridae). The two other most basal families (Lutodrilidae and Criodrilidae) are both much less diverse (with one and three species, respectively) and less explored by molecular phylogenetics approaches. Yet, their phylogenetic position and distribution (Nearctic and Palearctic, respectively) make them important pieces to understand the origin and paleobiogeography of this earthworm superfamily. It is well known that earthworm evolution closely reflects paleogeographic events [29,72], and the opening of the Atlantic Ocean would be expected to have determined the first stages of their diversification. Likewise, the formation of the Mediterranean Sea and the uplift of the Pyrenees and the Alps appear to have been key for the diversification of Hormogastridae and Lumbricidae. The origin of both can be found in the terrane, which consisted in the Northeastern Iberian Peninsula, Southern France, Corsica, and Sardinia [48,70]. Their latter eastward expansion, which led to the vast diversification of genera such as Dendrobaena, Eisenia, or Octodrilus, as well as the colonization of North America by the genera Eisenoides (Gates 1969) and Bimastos are comparatively poorly understood. In order to complete this relevant part of the evolutionary puzzle of Lumbricoidea, more accurate paleogeographic reconstructions of the Tertiary period and more complete sampling of the eastern Eurasian genera (including Perelia) are necessary.
6. Other Families: Remaining Questions
While important work has been conducted in other earthworm families (for example Rhinodrilidae (Benham 1890) [27,73], Megascolecidae (Rosa 1891) [41,45,74,75]), several families have received very little attention relative to the large percentage of earthworm diversity and occupied land masses they represent. Africa, South America, and North America display the strongest deficit of molecular phylogenetic research. Although molecular phylogenetic studies exist for most of those families, they often constitute isolated and unconnected attempts to answer very specific evolutionary questions.
Far from intending to diminish the value of those contributions, this work is meant to encourage earthworm experts to systematically apply available molecular phylogenetic tools in order to establish robust genus systems, spatio-temporal evolutionary frameworks (through biogeographical reconstructions and time-calibrated phylogenies), cryptic diversity assessment, and comparative phylogeographies. In cases where molecular biology facilities are not available, international collaboration should provide satisfactory solutions. However, the most advanced molecular phylogenetic tools will be moot if there are no taxonomists with expertise in the target taxa to guide them; the decline in the number of earthworm taxonomists in the last few decades is an alarming problem that affects all continents alike. Thus, the priority for active earthworm taxonomists should be to train a younger generation of researchers who could integrate knowledge of the intricate systematics of earthworms with the use of the rich molecular toolbox.
Author Contributions
Conceptualization, D.F.M., T.D., J.D. and M.N.; investigation, D.F.M., T.D., J.D. and M.N.; resources, D.F.M., T.D., J.D. and M.N.; writing—original draft preparation, D.F.M.; writing—review and editing, D.F.M., T.D., J.D. and M.N.; supervision, T.D., J.D. and M.N.; project administration, D.F.M., T.D., J.D. and M.N.; funding acquisition, D.F.M., T.D., J.D. and M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Xunta de Galicia (Consellería de Cultura, Educación e Ordenación Universitaria. Secretaria Xeral de Universidades under grant ED431B 2019/038), and by subproject Fauna Ibérica XII. Oligochaeta, Lumbricoidea: Lumbricidae, Hormogastridae #PGC2018-095851-B-C66 from the Spanish Ministry of Sciences, Innovation and Universities. DFM was funded by a Juan de La Cierva-Formación grant (FJCI-2017-32895) from the Spanish Ministry of Sciences, Innovation and Universities and by a Make Our Planet Great Again Postdoctoral grant from Campus France. MN was supported by Ramon y Cajal Fellowship (RYC2018-024654-I) from MCIN/AEI/10.13039/501100011033 and by “ESF: Investing in your future”.
Data Availability Statement
This manuscript did not produce new data; datasets were made available in the relevant publications.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pop, A.A.; Wink, M.; Pop, V.V. Use of 18S, 16S rDNA and cytochrome c oxidase sequences in earthworm taxonomy (Oligochaeta, Lumbricidae): The 7th international symposium on earthworm ecology·Cardiff·Wales·2002. Pedobiologia 2003, 47, 428–433. [Google Scholar] [CrossRef]
- Michaelsen, W. Oligochaeta. In Das Tierreich X.; Friedländer & Sohn: Berlin/Heidelberg, Germany, 1900; pp. 1–575. [Google Scholar]
- Pop, V. Zur phylogenie und Systematik der Lumbriciden. Zool. Jahrbücher Abt. Für Syst. Okol. Und Geogr. Der Tiere 1941, 74, 487–522. [Google Scholar]
- Omodeo, P. Contributo alla revisione dei Lumbricidae. Archivio Zoologico Italiano 1956, 41, 129–212. [Google Scholar]
- Gates, G.E. Contributions to a revision of the earthworm family Lumbricidae XII. Enterion mammale Savigny, 1826 and its position in the family. Megadrilogica 1975, 2, 1–5. [Google Scholar]
- Perel, T.S. A critical analysis of the system of family Lumbricidae. Zoologicheski Zhurnal 1976, 55, 823–836. (In Russian) [Google Scholar]
- Mršić, N. Monograph on Earthworms (Lumbricidae) of the Balkans I–II; Slovenska Akademija Znanosti in Umetnosti, Zazred za Naravoslovne Vede Opera. 31: Ljubljana, Slovenia, 1991; pp. 1–757. [Google Scholar]
- Qiu, J.P.; Bouché, M.B. Liste classée des taxons valides de Lombriciens (Oligochaeta: Lumbricoidea) apres l’etude des trios cinquieme d’entre-eux. Doc. Pédozoologiques Intégrologiques 1998, 4, 181–200. [Google Scholar]
- James, S.W.; Davidson, S.K. Molecular phylogeny of earthworms (Annelida: Crassiclitellata) based on 28S, 18S and 16S gene sequences. Invertebr. Syst. 2012, 26, 213–229. [Google Scholar] [CrossRef]
- Decaëns, T.; Porco, D.; Rougerie, R.; Brown, G.G.; James, S.W. Potential of DNA barcoding for earthworm research in taxonomy and ecology. Appl. Soil Ecol. 2013, 65, 35–42. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- King, R.A.; Tibble, A.L.; Symondson, W.O.C. Opening a can of worms: Unprecedented sympatric cryptic diversity within British lumbricid earthworms. Mol. Ecol. 2008, 17, 4684–4698. [Google Scholar] [CrossRef]
- Porco, D.; Chang, C.-H.; Dupont, L.; James, S.W.; Richard, B.; Decaëns, T. A reference library of DNA barcodes for the earthworms from Upper Normandy: Biodiversity assessment, new records, potential cases of cryptic diversity and ongoing speciation. Appl. Soil Ecol. 2018, 124, 362–371. [Google Scholar] [CrossRef]
- Novo, M.; Almodóvar, A.; Fernández, R.; Trigo, D.; Cosín, D.J.D. Cryptic speciation of hormogastrid earthworms revealed by mitochondrial and nuclear data. Mol. Phylogenet. Evol. 2010, 56, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Buckley, T.R.; James, S.; Allwood, J.; Bartlam, S.; Howitt, R.; Prada, D. Phylogenetic analysis of New Zealand earthworms (Oligochaeta: Megascolecidae) reveals ancient clades and cryptic taxonomic diversity. Mol. Phylogenetics Evol. 2011, 58, 85–96. [Google Scholar] [CrossRef]
- Marchán, D.F.; Hedde, M.; Lapied, E.; Maggia, M.E.; Novo, M.; Domínguez, J.; Decaëns, T. Contrasting phylogeographic patterns of earthworms (Crassiclitellata, Lumbricidae) on near-shore mediterranean islands. Eur. J. Soil Biol. 2020, 101, 103242. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Bloch, R.; Breinholt, J.W.; Pfenninger, M.; Domínguez, J. Taxonomic assessment of Lumbricidae (Oligochaeta) earthworm genera using DNA barcodes. Eur. J. Soil Biol. 2012, 48, 41–47. [Google Scholar] [CrossRef]
- Jamieson, B.G.; Tillier, S.; Tillier, A.; Justine, J.L.; Ling, E.; James, S.; McDonald, K.; Hugall, A.F. Phylogeny of the Megascolecidae and Crassiclitellata (Annelida, Oligochaeta): Combined versus partitioned analysis using nuclear (28S) and mitochondrial (12S, 16S) rDNA. Zoosystema 2002, 24, 707–734. [Google Scholar]
- Klarica, J.; Kloss-Brandstätter, A.; Traugott, M.; Juen, A. Comparing four mitochondrial genes in earthworms–implications for identification, phylogenetics, and discovery of cryptic species. Soil Biol. Biochem. 2012, 45, 23–30. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Simancas, B.; Cosín, D.J.D. Adding complexity to the complex: New insights into the phylogeny, diversification and origin of parthenogenesis in the Aporrectodea caliginosa species complex (Oligochaeta, Lumbricidae). Mol. Phylogenetics Evol. 2012, 64, 368–379. [Google Scholar] [CrossRef]
- Novo, M.; Almodovar, A.; Fernandez, R.; Trigo, D.; Díaz-Cosín, D.J.; Giribet, G. Appearances can be deceptive: Different diversification patterns within a group of Mediterranean earthworms (Oligochaeta, Hormogastridae). Mol. Ecol. 2012, 21, 3776–3793. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Porto, P.G.; Cosín, D.J.D.; Pérez-Losada, M. Multigene phylogeny reveals two new isolated and relic earthworm genera (Oligochaeta: Lumbricidae). Zool. J. Linn. Soc. 2017, 182, 258–274. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, J.; Dong, Y.; Qiu, J. Complete mitochondrial genome of four pheretimoid earthworms (Clitellata: Oligochaeta) and their phylogenetic reconstruction. Gene 2015, 574, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Dupont, L.; Porco, D.; Symondson, W.O.C.; Roy, V. Hybridization relics complicate barcode-based identification of species in earthworms. Mol. Ecol. Resour. 2016, 16, 883–894. [Google Scholar] [CrossRef]
- Taheri, S.; James, S.; Roy, V.; Decaëns, T.; Williams, B.W.; Anderson, F.; Rougerie, R.; Chang, C.H.; Brown, G.; Cunha, L.; et al. Complex taxonomy of the ‘brush tail’peregrine earthworm Pontoscolex corethrurus. Mol. Phylogenet. Evol. 2018, 124, 60–70. [Google Scholar] [CrossRef]
- Novo, M.; Fernández, R.; Andrade, S.C.; Marchán, D.F.; Cunha, L.; Cosín, D.J.D. Phylogenomic analyses of a Mediterranean earthworm family (Annelida: Hormogastridae). Mol. Phylogenet. Evol. 2016, 94, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.E.; Williams, B.W.; Horn, K.M.; Erséus, C.; Halanych, K.M.; Santos, S.R.; James, S.W. Phylogenomic analyses of Crassiclitellata support major Northern and Southern Hemisphere clades and a Pangaean origin for earthworms. BMC Evol. Biol. 2017, 17, 123. [Google Scholar]
- Faircloth, B.C.; Glenn, T.C. Not All Sequence Tags Are Created Equal: Designing and Validating Sequence Identification Tags Robust to Indels. PLoS ONE 2012, 7, e42543. [Google Scholar] [CrossRef]
- Lemmon, A.R.; Emme, S.A.; Lemmon, E.M. Anchored Hybrid Enrichment for Massively High-Throughput Phylogenomics. Syst. Biol. 2012, 61, 727–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, B.R.; Gamble, T.; Jackman, T.R. Optimizing Phylogenomics with Rapidly Evolving Long Exons: Comparison with Anchored Hybrid Enrichment and Ultraconserved Elements. Mol. Biol. Evol. 2020, 37, 904–922. [Google Scholar] [CrossRef]
- Crawford, N.G.; Parham, J.F.; Sellas, A.B.; Faircloth, B.C.; Glenn, T.C.; Papenfuss, T.J.; Henderson, J.B.; Hansen, M.H.; Simison, W.B. A phylogenomic analysis of turtles. Mol. Phylogenet. Evol. 2015, 83, 250–257. [Google Scholar] [CrossRef]
- Streicher, J.W.; Wiens, J.J. Phylogenomic analyses of more than 4000 nuclear loci resolve the origin of snakes among lizard families. Biol. Lett. 2017, 13, 20170393. [Google Scholar] [CrossRef]
- Phillips, A.J.; Dornburg, A.; Zapfe, K.L.; Anderson, F.E.; James, S.W.; Erséus, C.; Lemmon, E.M.; Lemmon, A.R.; Williams, B.W. Phylogenomic Analysis of a Putative Missing Link Sparks Reinterpretation of Leech Evolution. Genome Biol. Evol. 2019, 11, 3082–3093. [Google Scholar] [CrossRef] [Green Version]
- Emerson, K.J.; Merz, C.R.; Catchen, J.M.; Hohenlohe, P.A.; Cresko, W.A.; Bradshaw, W.E.; Holzapfel, C.M. Resolving postglacial phylogeography using high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 16196–16200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.; Cunha, L.; Sechi, P.; Kille, P.; Spurgeon, D. Genetic variation in populations of the earthworm, Lumbricus rubellus, across contaminated mine sites. BMC Genet. 2017, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Marchán, D.F.; Novo, M.; Sánchez, N.; Domínguez, J.; Cosín, D.J.D.; Fernandez, R. Local adaptation fuels cryptic speciation in terrestrial annelids. Mol. Phylogenetics Evol. 2020, 146, 106767. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; Domínguez, J.; Díaz Cosín, D.J.; Novo, M. Genome-informed integrative taxonomic description of three cryptic species in the earthworm genus Carpetania (Oligochaeta, Hormogastridae). Syst. Biodivers. 2020, 18, 203–215. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, J.; Dong, Y.; Zhao, Q.; Sun, J.; Qiu, J. Unearthing the genetic divergence and gene flow of the earthworm Amynthas_YN2017 sp. (Oligochaeta: Megascolecidae) populations based on restriction site-associated DNA sequencing. Eur. J. Soil Biol. 2020, 99, 103210. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Marchan, D.F.; Fernández, R.; de Sosa, I.; Cosin, D.J.D.; Novo, M. Pinpointing cryptic borders: Fine-scale phylogeography and genetic landscape analysis of the Hormogaster elisae complex (Oligochaeta, Hormogastridae). Mol. Phylogenet. Evol. 2017, 112, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Almodóvar, A.; Fernández, R.; Giribet, G.; Cosín, D.J.D. Understanding the biogeography of a group of earthworms in the Mediterranean basin—The phylogenetic puzzle of Hormogastridae (Clitellata: Oligochaeta). Mol. Phylogenet. Evol. 2011, 61, 125–135. [Google Scholar] [CrossRef]
- Shen, H.-P.; Chang, C.-H.; Ota, H. The biogeographical history of giant earthworms of the Metaphire formosae species group (Clitellata: Megascolecidae) in Taiwan and the Ryukyu Archipelago, with the description of a new species from Yonagunijima, Southern Ryukyus. Org. Divers. Evol. 2021. [Google Scholar] [CrossRef]
- Zhao, Q.; Cluzeau, D.; Jiang, J.; Petit, E.J.; Briard, C.; Sun, J.; Prinzing, A.; Qiu, J. Molecular Phylogeny of Pheretimoid Earthworms (Haplotaxina: Megascolecidae) Based on Mitochondrial DNA in Hainan Island, China. Mol. Biol. 2015, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- De Sosa, I.; Cosín, D.J.D.; Csuzdi, C.; Paoletti, M.G.; Marchán, D.F. Placing Eophila tellinii (Oligochaeta, Lumbricidae) in a molecular phylogenetic context advances the century-old controversy around the problematic genus. Eur. J. Soil Biol. 2019, 94, 103114. [Google Scholar] [CrossRef]
- Marchán, D.F.; Csuzdi, C.; Decaëns, T.; Szederjesi, T.; Pizl, V.; Domínguez, J. The disjunct distribution of relict earthworm genera clarifies the early historical biogeography of the Lumbricidae (Crassiclitellata, Annelida). J. Zoöl. Syst. Evol. Res. 2021, 59, 1703–1717. [Google Scholar] [CrossRef]
- Chen, D.; Hosner, P.A.; Dittmann, D.L.; O’Neill, J.P.; Birks, S.M.; Braun, E.L.; Kimball, R.T. Divergence time estimation of Galliformes based on the best gene shopping scheme of ultraconserved elements. BMC Ecol. Evol. 2021, 21, 209. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Breinholt, J.W.; Stojanovic, M.; James, S.W.; Pérez-Losada, M. Underground evolution: New roots for the old tree of lumbricid earthworms. Mol Phylogenet Evol. 2015, 83, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Jiménez Pinadero, S.; Marchán, D.F.; Novo, M.; Trigo, D.; Domínguez, J.; Cosín, D.J.D. Sorry atlanticus, you are not my type: Molecular assessment splits Zophoscolex (Lumbricidae: Crassiclitellata) into French and Iberian genera. Zool. J. Linn. Soc. 2021. [Google Scholar] [CrossRef]
- Marchán, D.F.; Jiménez, S.; Decaëns, T.; Domínguez, J. Systematic revision of Gatesona (Crassiclitellata, Lumbricidae), an endemic earthworm genus from the Massif Central (France). PLoS ONE 2021, 16, e0255978. [Google Scholar] [CrossRef]
- Csuzdi, C.; Chang, C.-H.; Pavlícek, T.; Szederjesi, T.; Esopi, D.; Szlavecz, K. Molecular phylogeny and systematics of native North American lumbricid earthworms (Clitellata: Megadrili). PLoS ONE 2017, 12, e0181504. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Losada, M.; Ricoy, M.; Marshall, J.C.; Domínguez, J. Phylogenetic assessment of the earthworm Aporrectodea caliginosa species complex (Oligochaeta: Lumbricidae) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 52, 293–302. [Google Scholar] [CrossRef]
- Fernández, R.; Novo, M.; Marchán, D.F.; Cosín, D.J.D. Diversification patterns in cosmopolitan earthworms: Similar mode but different tempo. Mol. Phylogenet. Evol. 2016, 94, 701–708. [Google Scholar] [CrossRef]
- Dupont, L.; Lazrek, F.; Porco, D.; King, R.; Rougerie, R.; Symondson, W.; Livet, A.; Richard, B.; Decaëns, T.; Butt, K.; et al. New insight into the genetic structure of the Allolobophora chlorotica aggregate in Europe using microsatellite and mitochondrial data. Pedobiologia 2011, 54, 217–224. [Google Scholar] [CrossRef]
- Marchán, D.F.; Cosín, D.J.D.; Novo, M. Why are we blind to cryptic species? Lessons from the eyeless. Eur. J. Soil Biol. 2018, 86, 49–51. [Google Scholar] [CrossRef]
- Szederjesi, T.; Pop, V.V.; Pavlíček, T.; MÁrton, O.; KrÍzsik, V.; Csuzdi, C. Integrated taxonomy reveals multiple species in the Dendrobaena byblica (Rosa, 1893) complex (Oligochaeta: Lumbricidae). Zool. J. Linn. Soc. 2018, 182, 500–516. [Google Scholar] [CrossRef]
- Martinsson, S.; Klinth, M.; Erséus, C. Distribution and genetic diversity of two earthworms, Helodrilus oculatus and Satchellius mammalis (Clitellata: Lumbricidae) in Scandinavia. Fauna Nor. 2021, 41, 1–14. [Google Scholar] [CrossRef]
- Marchán, D.F.; Decaëns, T.; Cosín, D.J.D.; Hedde, M.; Lapied, E.; Domínguez, J. French Mediterranean islands as a refuge of relic earthworm species: Cataladrilus porquerollensis sp. nov. and Scherotheca portcrosana sp. nov. (Crassiclitellata, Lumbricidae). Eur. J. Taxon. 2020, 701, 1–22. [Google Scholar] [CrossRef]
- Bache, F.; Olivet, J.L.; Gorini, C.; Aslanian, D.; Labails, C.; Rabineau, M. Evolution of rifted continental margins: The case of the Gulf of Lions (Western Mediterranean Basin). Earth Planet. Sci. Lett. 2010, 292, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Cosin, D.D.; Novo, M.; Fernández, R.; Marchán, D.F.; Gutiérrez, M. A new earthworm species within a controversial genus: Eiseniona gerardoi sp. n. (Annelida, Lumbricidae)—Description based on morphological and molecular data. ZooKeys 2014, 399, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Novo, M.; Fernández, R.; Marchán, D.F.; Gutiérrez, M.; Cosín, D.J.D. Compilation of morphological and molecular data, a necessity for taxonomy: The case of Hormogaster abbatissae sp. n.(Annelida, Clitellata, Hormogastridae). ZooKeys 2012, 242, 1. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; Novo, M.; Cosin, D.D. New light into the hormogastrid riddle: Morphological and molecular description of Hormogaster joseantonioi sp. n. (Annelida, Clitellata, Hormogastridae). ZooKeys 2014, 414, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rota, E.; Marchan, D.F.; Omodeo, P. Hormogaster regina sp. n. (Annelida: Clitellata: Hormogastridae): A giant earthworm from Spanish Catalonia, described from morphological and molecular evidence. Zool. Anz. A J. Comp. Zool. 2016, 261, 56–65. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernandez, R.; Sánchez, N.; De Sosa, I.; Cosín, D.J.D.; Novo, M. Insights into the diversity of Hormogastridae (Annelida, Oligochaeta) with descriptions of six new species. Zootaxa 2018, 4496, 65–95. [Google Scholar] [CrossRef]
- Marchan, D.F.; Novo, M.; Fernández, R.; De Sosa, I.; Trigo, D.; Cosín, D.J.D. Evaluating evolutionary pressures and phylogenetic signal in earthworms: A case study—The number of typhlosole lamellae in Hormogastridae (Annelida, Oligochaeta). Zool. J. Linn. Soc. 2016, 178, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Marchán, D.F.; Sánchez, N.; Novo, M.; Fernández, R.; Pardos, F.; Cosín, D.J.D. Cryptic characters for cryptic taxa: On the taxonomic utility of the genital chaetae in earthworms (Oligochaeta, Hormogastridae). Zool. Anz. A J. Comp. Zool. 2016, 264, 17–28. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; de Sosa, I.; Sánchez, N.; Cosín, D.J.D.; Novo, M. Integrative systematic revision of a Mediterranean earthworm family: Hormogastridae (Annelida, Oligochaeta). Invertebr. Syst. 2018, 32, 652–671. [Google Scholar] [CrossRef]
- Novo, M.; Fernández, R.; Fernandez Marchan, D.; Trigo, D.; Diaz Cosin, D.J.; Giribet, G. Unearthing the historical biogeography of Mediterranean earthworms (Annelida: Hormogastridae). J. Biogeogr. 2015, 42, 751–762. [Google Scholar] [CrossRef]
- Tilikj, N.; Novo, M. How to resist soil desiccation: Transcriptional changes in a Mediterranean earthworm during aestivation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 264, 111112. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Gutiérrez, M.; Cosín, D.J.D. Earthworms, good indicators for palaeogeographical studies? Testing the genetic structure and demographic history in the peregrine earthworm Aporrectodea trapezoides (Dugès, 1828) in southern Europe. Soil Biol. Biochem. 2013, 58, 127–135. [Google Scholar] [CrossRef]
- Taheri, S.; Decaëns, T.; Cunha, L.; Brown, G.G.; Da Silva, E.; Bartz, M.; Baretta, D.; Dupont, L. Genetic evidence of multiple introductions and mixed reproductive strategy in the peregrine earthworm Pontoscolex corethrurus. Biol. Invasions 2020, 22, 2545–2557. [Google Scholar] [CrossRef]
- Aspe, N.M.; James, S.W. Molecular phylogeny and biogeographic distribution of pheretimoid earthworms (clitellata: Megascolecidae) of the Philippine archipelago. Eur. J. Soil Biol. 2018, 85, 89–97. [Google Scholar] [CrossRef]
- Jiang, J.; Qiu, J. Origin and evolution of earthworms belonging to the family Megascolecidae in China. Biodivers. Sci. 2018, 26, 1074–1082. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Main molecular phylogenetic techniques available for earthworm systematics and evolutionary research.
Figure 1.
Main molecular phylogenetic techniques available for earthworm systematics and evolutionary research.

Figure 2.
Modified from [48]. Bayesian phylogenetic inference based on the concatenated sequences of the molecular markers COI, COII, 16S, tRNAs (Ala, Leu), 12S, ND1, and 28S. Clades referenced in the main text are highlighted in different colours; clades for which significant advances have been achieved are shown in green; clades for which remaining questions exist are shown in orange.
Figure 2.
Modified from [48]. Bayesian phylogenetic inference based on the concatenated sequences of the molecular markers COI, COII, 16S, tRNAs (Ala, Leu), 12S, ND1, and 28S. Clades referenced in the main text are highlighted in different colours; clades for which significant advances have been achieved are shown in green; clades for which remaining questions exist are shown in orange.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marchán, D.F.; Decaëns, T.; Domínguez, J.; Novo, M. Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions. Diversity 2022, 14, 30. https://doi.org/10.3390/d14010030
AMA Style
Marchán DF, Decaëns T, Domínguez J, Novo M. Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions. Diversity. 2022; 14(1):30. https://doi.org/10.3390/d14010030
Chicago/Turabian StyleMarchán, Daniel Fernández, Thibaud Decaëns, Jorge Domínguez, and Marta Novo. 2022. "Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions" Diversity 14, no. 1: 30. https://doi.org/10.3390/d14010030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.