
Toxicogenomics of the Freshwater Oligochaete, Tubifex tubifex (Annelida, Clitellata), in Acute Water-Only Exposure to Arsenic
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Water Acute Toxicity Test for As
2.2. Reference Transcriptome
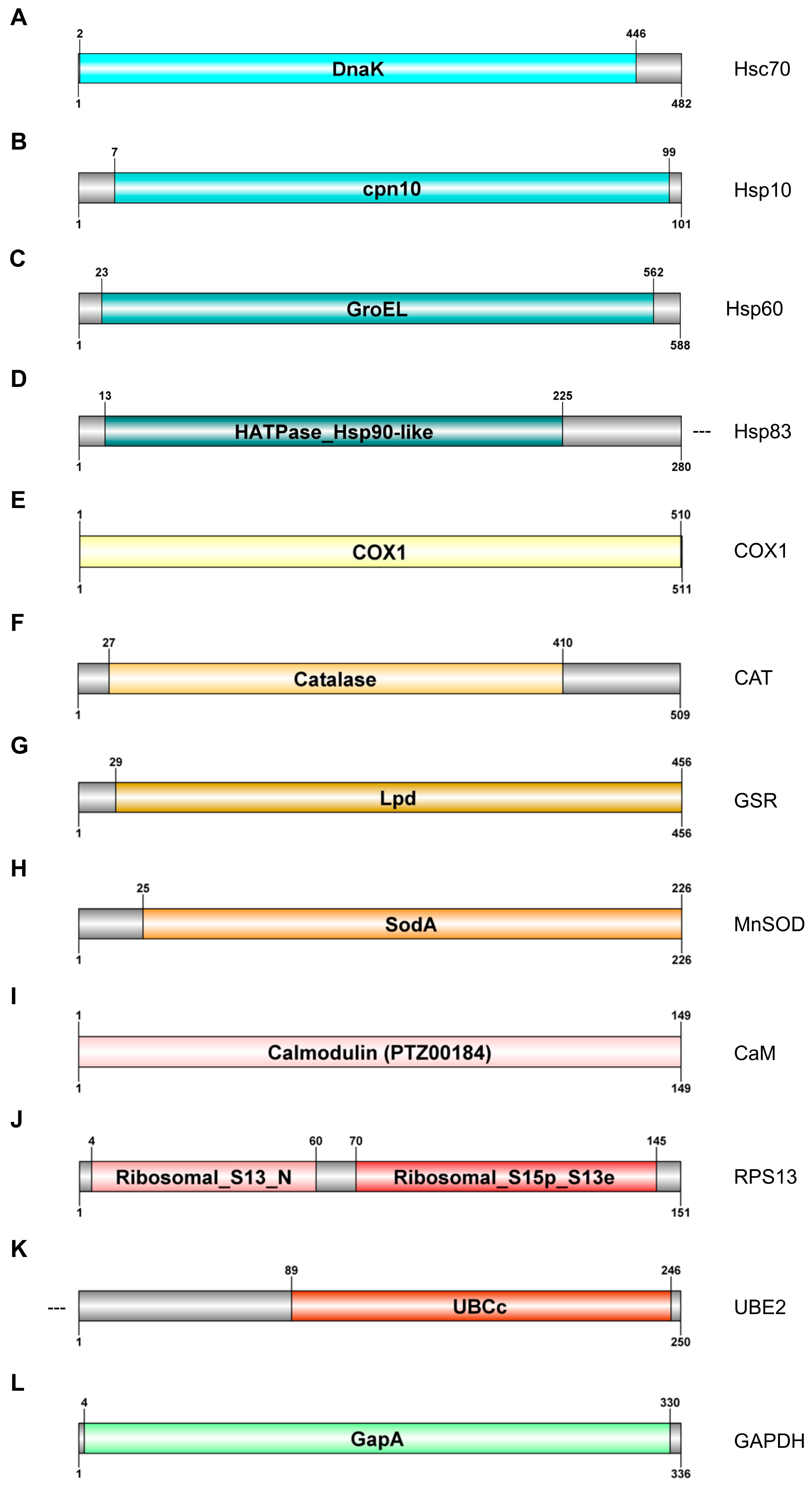
2.3. Gene Characterisation
2.3.1. Cell Stress Response Genes
2.3.2. Energy Metabolism and Oxidative Stress Response
2.3.3. Homeostasis Maintenance
2.3.4. Reference Genes
2.4. Gene Expression Study
2.4.1. Cell Stress Response
2.4.2. Energy Metabolism and Oxidative Stress Biomarkers
2.4.3. Transcriptional Alterations in Homeostasis-Related Biomarkers
3. Discussion
4. Materials and Methods
4.1. Water-Only Acute Toxicity Test for As
4.2. Gene Expression Study
4.2.1. Sequencing, De Novo Assembly, and Annotation of a Reference Transcriptome
4.2.2. Gene Characterisation
4.2.3. RNA Extraction
4.2.4. Gene Expression Analysis
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Arsenic (mg L−1) | Temperature (°C) | pH | Conductivity (µS cm−1) | Dissolved Oxygen (% SAT) | Dissolved Oxygen (mg L−1) |
|---|---|---|---|---|---|
| 0 | 23.6 | 7.76 | 319 | 85.8 | 7.18 |
| 38 | 23.5 | 8.12 | 415 | 85.2 | 7.12 |
| 63 | 23.5 | 8.25 | 477 | 85.4 | 7.09 |
| 100 | 23.7 | 8.28 | 577 | 86.1 | 7.19 |
| 159 | 23.5 | 8.31 | 733 | 84.0 | 7.03 |
| 250 | 23.6 | 8.40 | 979 | 85.2 | 7.10 |
| 407 | 23.7 | 8.47 | 1377 | 86.7 | 7.21 |
| Gene | Species | Accession Number | Length (aa) | Identity (%) |
|---|---|---|---|---|
| Hsc70 | Tubifex tubifex | PP213149 | 482 | - |
| Biomphalaria glabrata | XP_013082114.1 | 649 | 89 | |
| Sacostrea echinata | XP_061181462.1 | 659 | 89 | |
| Hsp10 | Tubifex tubifex | PP213150 | 101 | - |
| Crassostrea gigas | XP_011456448.1 | 101 | 76 | |
| Physella acuta | XP_059140133.1 | 101 | 75 | |
| Hsp60 | Tubifex tubifex | PP213151 | 588 | - |
| Helobdella robusta | XP_009028388.1 | 584 | 78 | |
| Eisenia fetida | WRM33298.1 | 586 | 80 | |
| Hsp83 | Tubifex tubifex | PP213152 | 280 * | - |
| Halyomorpha halys | XP_014284945.1 | 723 | 87 | |
| Arma chinensis | WGV41501.1 | 723 | 82 | |
| COX1 | Tubifex tubifex | PP212898 | 511 | - |
| Limnodrilus hoffmeisteri | QWT71538.1 | 512 | 86 | |
| Nais communis | QUT13265.1 | 512 | 81 | |
| Cat | Tubifex tubifex | PP213153 | 509 | - |
| Eisenia fetida | AEO50756.2 | 505 | 74 | |
| Haliotis madaka | ALU63753.1 | 509 | 75 | |
| GSR | Tubifex tubifex | PP213154 | 456 | - |
| Haliotis fulgens | AXR98615.1 | 452 | 65 | |
| Pomacea canaliculata | XP_025078581.1 | 452 | 65 | |
| MnSOD | Tubifex tubifex | PP213155 | 226 | - |
| Eisenia andrei | AMZ80133.2 | 225 | 69 | |
| Zootermopsis nevadensis | XP_021916368.1 | 225 | 69 | |
| CaM | Tubifex tubifex | PP213156 | 149 | - |
| Lumbricus rubellus | Q9GRJ1.3 | 149 | 100 | |
| Pinctada fucata | AAQ20043.1 | 149 | 99 | |
| RpS13 | Tubifex tubifex | PP213157 | 151 | - |
| Arenicola marina | ABW23143.1 | 151 | 91 | |
| Lumbricus rubellus | O77303.3 | 151 | 88 | |
| UBE2 | Tubifex tubifex | PP213158 | 250 * | - |
| Branchiostoma floridae | XP_035678611.1 | 342 | 65 | |
| Lingula anatina | XP_013421956.1 | 319 | 63 | |
| Gapdh | Tubifex tubifex | PP213159 | 336 | - |
| Enchytraeus albidus | AOR07119.1 | 336 | 81 | |
| Mesenchytraeus hydrius | AOR07125.1 | 336 | 78 |
References
- Eisler, R. Handbook of Chemical Risk Assesment: Health Hazards to Humans, Plants, and Animals. Vol 1: Metals; Volume 2: Organics; Volume 3: Metalloids, Radiation, Cumulative Index to Chemicals, and Species By Ronald Eisler (Patuxent Wildlife Research Center). J. Am. Chem. Soc. 2000, 122, 12067. [Google Scholar]
- Loredo, J.; Ordóñez, A.; Alvarez, R. Environmental impact of toxic metals and metalloids from the Munon Cimero mercury-mining area (Asturias, Spain). J. Hazard. Mater. 2006, 136, 455–467. [Google Scholar] [CrossRef]
- Méndez-Fernández, L.; Rodriguez, P.; Martínez-Madrid, M. Sediment toxicity and bioaccumulation assessment in abandoned copper and mercury mining areas of the Nalón River basin (Spain). Arch. Environ. Contam. Toxicol. 2015, 68, 107–123. [Google Scholar] [CrossRef]
- Rainbow, P.S. Trace Metals in the Environment and Living Organisms: The British Isles as a Case Study; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Bhattacharya, P.; Welch, A.H.; Stollenwerk, K.G.; McLaughlin, M.J.; Bundschuh, J.; Panaullah, G. Arsenic in the environment: Biology and chemistry. Sci. Total Environ. 2007, 379, 109–120. [Google Scholar] [CrossRef]
- Rodríguez, P.; Moreno-Ocio, I.; Martínez-Madrid, M.; Costas, N.; Pardo, I.; Méndez-Fernández, L. Proposal of integrative scores and biomonitor selection for metal bioaccumulation risk assessment in mine-impacted rivers. Aquat. Toxicol. 2021, 238, 105918. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Ocio, I.; Méndez-Fernández, L.; Martínez-Madrid, M.; Costas, N.; Pardo, I.; Rodriguez, P. Developing As and Cu Tissue Residue Thresholds to Attain the Good Ecological Status of Rivers in Mining Areas. Arch. Environ. Contam. Toxicol. 2022, 82, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The role of biomarkers in the assessment of aquatic ecosystem health. Integr. Environ. Assess. Manag. 2014, 10, 327–341. [Google Scholar] [CrossRef]
- Colin, N.; Porte, C.; Fernandes, D.; Barata, C.; Padrós, F.; Carrassón, M.; Monroy, M.; Cano-Rocabayera, O.; de Sostoa, A.; Piña, B.; et al. Ecological relevance of biomarkers in monitoring studies of macro-invertebrates and fish in Mediterranean rivers. Sci. Total Environ. 2016, 540, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Minatel, B.C.; Sage, A.P.; Anderson, C.; Hubaux, R.; Marshall, E.A.; Lam, W.L.; Martinez, V.D. Environmental arsenic exposure: From genetic susceptibility to pathogenesis. Environ. Int. 2018, 112, 183–197. [Google Scholar] [CrossRef]
- Hubaux, R.; Becker-Santos, D.D.; Enfield, K.S.; Rowbotham, D.; Lam, S.; Lam, W.L.; Martinez, V.D. Molecular features in arsenic-induced lung tumors. Mol. Cancer 2013, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Mermillod-Blondin, F.; Nogaro, G.; Datry, T.; Malard, F.; Gibert, J. Do tubificid worms influence the fate of organic matter and pollutants in stormwater sediments? Environ. Pollut. 2005, 134, 57–69. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Lemoine, D. Ecosystem engineering by tubificid worms stimulates macrophyte growth in poorly oxygenated wetland sediments. Funct. Ecol. 2010, 24, 444–453. [Google Scholar] [CrossRef]
- Przeslawski, R.; Zhu, Q.; Aller, R. Effects of abiotic stressors on infaunal burrowing and associated sediment characteristics. Mar. Ecol.-Prog. Ser. 2009, 392, 33–42. [Google Scholar] [CrossRef]
- Lagauzère, S.; Terrail, R.; Bonzom, J.M. Ecotoxicity of uranium to Tubifex tubifex worms (Annelida, Clitellata, Tubificidae) exposed to contaminated sediment. Ecotoxicol. Environ. Saf. 2009, 72, 527–537. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Foulquier, A.; Gilbert, F.; Navel, S.; Montuelle, B.; Bellvert, F.; Comte, G.; Grossi, V.; Fourel, F.; Lecuyer, C.; et al. Benzo(a)pyrene inhibits the role of the bioturbator Tubifex tubifex in river sediment biogeochemistry. Sci. Total Environ. 2013, 450–451, 230–241. [Google Scholar] [CrossRef]
- Nie, X.B.; Li, Z.H.; Long, Y.N.; He, P.P.; Xu, C. Chlorine inactivation of Tubifex tubifex in drinking water and the synergistic effect of sequential inactivation with UV irradiation and chlorine. Chemosphere 2017, 177, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, P.; Reynoldson, T.B. The Pollution Biology of Aquatic Oligochaetes; Springer Science Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Chapman, P.M. Utility and relevance of aquatic oligochaetes in ecological risk assessment. In Aquatic Oligochaete Biology VIII; Springer: Dordrecht, The Netherlands, 2001; pp. 149–169. [Google Scholar]
- ASTM E1706-05; Standard Test Method for Measuring the Toxicity of Sediment-Associated Contaminants with Freshwater Invertebrates. American Society for Testing and Materials (ASTM): Philadelphia, PA, USA, 2005.
- Organization for Economic Cooperation Development (OECD). Test No. 315: Guidelines for the Testing of Chemicals: Bioaccumulation in Sediment-Dwelling Benthic Oligochaetes; OECD Publishing: Paris, France, 2008; p. 33. [Google Scholar]
- Lobo, H.; Méndez-Fernández, L.; Martínez-Madrid, M.; Daam, M.A.; Espíndola, E.L. Acute toxicity of zinc and arsenic to the warmwater aquatic oligochaete Branchiura sowerbyi as compared to its coldwater counterpart Tubifex tubifex (Annelida, Clitellata). J. Soils Sediments 2016, 16, 2766–2774. [Google Scholar] [CrossRef]
- Lobo, H.; Méndez-Fernández, L.; Martínez-Madrid, M.; Rodriguez, P.; Daam, M.A.; Espíndola, E.L. Bioaccumulation and chronic toxicity of arsenic and zinc in the aquatic oligochaetes Branchiura sowerbyi and Tubifex tubifex (Annelida, Clitellata). Aquat. Toxicol. 2021, 239, 105955. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Lahive, E.; Díez Ortiz, M.; Spurgeon, D.J.; Kille, P. Toxicogenomics in a soil sentinel exposure to Zn nanoparticles and ions reveals the comparative role of toxicokinetic and toxicodynamic mechanisms. Environ. Sci. Nano 2020, 7, 1464–1480. [Google Scholar] [CrossRef]
- de Sosa, I.; Verdes, A.; Tilikj, N.; Marchán, D.F.; Planelló, R.; Herrero, Ó.; Almodóvar, A.; Cosín, D.D.; Novo, M. How to thrive in unstable environments: Gene expression profile of a riparian earthworm under abiotic stress. Sci. Total Environ. 2022, 817, 152749. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Chatterjee, A.; Chatterjee, S.; Saha, N.C. Oxidative stress in benthic oligochaete worm, Tubifex tubifex induced by sublethal exposure to a cationic surfactant cetylpyridinium chloride and an anionic surfactant sodium dodecyl sulfate. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2021, 240, 108906. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Chatterjee, A.; Chatterjee, S.; Saha, N.C. Acute toxicity and sublethal effects of sodium laureth sulfate on oxidative stress enzymes in benthic oligochaete worm, Tubifex tubifex. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 243, 108998. [Google Scholar] [CrossRef]
- Petrusevski, B.; Sharma, S.; Schippers, J.C.; Shordt, K. Arsenic in Drinking Water; IRC International Water and Sanitation Centre: Delft, The Netherlands, 2007; Volume 17, pp. 36–44. [Google Scholar]
- Liu, T.; Zhang, L. Multigenerational effects of arsenate on development and reproduction in marine copepod Tigriopus japonicus. Chemosphere 2023, 342, 140158. [Google Scholar] [CrossRef]
- Eisler, R. Arsenic Hazards to Humans, Plants, and Animals from Gold Mining. Rev. Environ. Contam. Toxicol. 2004, 180, 133–165. [Google Scholar] [PubMed]
- Zhang, X.; Zhong, H.Q.; Chu, Z.W.; Zuo, X.; Wang, L.; Ren, X.L.; Ma, H.; Du, R.Y.; Ju, J.J.; Ye, X.L.; et al. Arsenic induces transgenerational behavior disorders in Caenorhabditis elegans and its underlying mechanisms. Chemosphere 2020, 252, 126510. [Google Scholar] [CrossRef]
- Fargasova, A. Toxicity of metals on Daphnia magna and Tubifex tubifex. Ecotoxicol. Environ. Saf. 1994, 27, 210–213. [Google Scholar] [CrossRef]
- Khangarot, B.S. Toxicity of metals to a freshwater tubificid worm Tubifex tubifex (Muller). Bull. Environ. Contam. Toxicol. 1991, 46, 906–912. [Google Scholar] [CrossRef]
- Haque, M.N.; Nam, S.E.; Kim, B.M.; Kim, K.; Rhee, J.S. Temperature elevation stage-specifically increases metal toxicity through bioconcentration and impairment of antioxidant defense systems in juvenile and adult marine mysids. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2020, 237, 108831. [Google Scholar] [CrossRef] [PubMed]
- Rathore, R.S.; Khangarot, B.S. Effects of temperature on the sensitivity of sludge worm Tubifex tubifex Müller to selected heavy metals. Ecotoxicol. Environ. Saf. 2002, 53, 27–36. [Google Scholar] [CrossRef]
- Wang, W. Factors affecting metal toxicity to (and accumulation by) aquatic organisms—Overview. Environ. Int. 1987, 13, 437–457. [Google Scholar] [CrossRef]
- Li, D.; Wei, Y.; Xu, S.; Niu, Q.; Zhang, M.; Li, S.; Jing, M. A systematic review and meta-analysis of bidirectional effect of arsenic on ERK signaling pathway. Mol. Med. Rep. 2018, 17, 4422–4432. [Google Scholar] [CrossRef] [PubMed]
- Jou, Y.C.; Wang, S.C.; Dai, Y.C.; Chen, S.Y.; Shen, C.H.; Lee, Y.R.; Chen, L.C.; Liu, Y.W. Gene expression and DNA methylation regulation of arsenic in mouse bladder tissues and in human urothelial cells. Oncol. Rep. 2019, 42, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Xia, Y.; Wade, T.J.; DeMarini, D.M.; Davidson, M.; Mumford, J. Altered gene expression by low-dose arsenic exposure in humans and cultured cardiomyocytes: Assessment by real-time PCR arrays. Int. J. Environ. Res. Public Health 2011, 8, 2090–2108. [Google Scholar] [CrossRef]
- Rahaman, M.S.; Yamasaki, S.; Binte Hossain, K.F.; Hosokawa, T.; Saito, T.; Kurasaki, M. Effects of curcumin, D-pinitol alone or in combination in cytotoxicity induced by arsenic in PC12 cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 144, 111577. [Google Scholar] [CrossRef]
- Susan, A.; Rajendran, K.; Sathyasivam, K.; Krishnan, U.M. An overview of plant-based interventions to ameliorate arsenic toxicity. Biomed. Pharmacother. 2019, 109, 838–852. [Google Scholar] [CrossRef]
- Anushree, A.M.Z.; Bilgrami, A.L.; Ahsan, J. Acute Exposure to Arsenic Affects Pupal Development and Neurological Functions in Drosophila melanogaster. Toxics 2023, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Mangu, J.C.K.; Rai, N.; Mandal, A.; Olsson, P.E.; Jass, J. Lysinibacillus sphaericus mediates stress responses and attenuates arsenic toxicity in Caenorhabditis elegans. Sci. Total Environ. 2022, 835, 155377. [Google Scholar] [CrossRef]
- Im, J.; Chatterjee, N.; Choi, J. Genetic, epigenetic, and developmental toxicity of Chironomus riparius raised in metal-contaminated field sediments: A multi-generational study with arsenic as a second challenge. Sci. Total Environ. 2019, 672, 789–797. [Google Scholar] [CrossRef]
- Frat, L.; Chertemps, T.; Pesce, E.; Bozzolan, F.; Dacher, M.; Planelló, R.; Herrero, O.; Llorente, L.; Moers, D.; Siaussat, D. Single and mixed exposure to cadmium and mercury in Drosophila melanogaster: Molecular responses and impact on post-embryonic development. Ecotoxicol. Environ. Saf. 2021, 220, 112377. [Google Scholar] [CrossRef]
- Xiao, X.; Tong, Y.; Wang, D.; Gong, Y.; Zhou, Z.; Liu, Y.; Huang, H.; Zhang, B.; Li, H.; You, J. Spatial distribution of benthic toxicity and sediment-bound metals and arsenic in Guangzhou urban waterways: Influence of land use. J. Hazard. Mater. 2022, 439, 129634. [Google Scholar] [CrossRef]
- Nadeau, D.; Corneau, S.; Plante, I.; Morrow, G.; Tanguay, R.M. Evaluation for Hsp70 as a biomarker of effect of pollutants on the earthworm Lumbricus terrestris. Cell Stress Chaperones 2001, 6, 153. [Google Scholar] [CrossRef]
- Homa, J.; Olchawa, E.; Stürzenbaum, S.R.; Morgan, A.J.; Plytycz, B. Early-phase immunodetection of metallothionein and heat shock proteins in extruded earthworm coelomocytes after dermal exposure to metal ions. Environ. Pollut. 2005, 135, 275–280. [Google Scholar] [CrossRef]
- Wen, F.; Yang, J.; Huang, X.; Huang, X. Analysis of Differential Gene Expression of the Aquatic Insect Protohermes costalis (Walker) (Megaloptera: Corydalidae) in Response to Cadmium Exposure. Environ. Entomol. 2022, 51, 815–823. [Google Scholar] [CrossRef]
- Jeyachandran, S.; Chellapandian, H.; Park, K.; Kwak, I.S. A Review on the Involvement of Heat Shock Proteins (Extrinsic Chaperones) in Response to Stress Conditions in Aquatic Organisms. Antioxidants 2023, 12, 1444. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Avadhani, N.G. Cytochrome c oxidase dysfunction in oxidative stress. Free Radic. Biol. Med. 2012, 53, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.A.; Schmidt, B.; Hiser, C.; Westley, E.; Ferguson-Miller, S. Membrane Potential-controlled Inhibition of Cytochrome c Oxidase by Zinc. J. Biol. Chem. 2002, 277, 14894–14901. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Severson, P.; Pacheco, S.; Futscher, B.W.; Klimecki, W.T. Arsenic exposure induces the Warburg effect in cultured human cells. Toxicol. Appl. Pharmacol. 2013, 271, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Luz, A.L.; Godebo, T.R.; Bhatt, D.P.; Ilkayeva, O.R.; Maurer, L.L.; Hirschey, M.D.; Meyer, J.N. From the cover: Arsenite uncouples mitochondrial respiration and induces a Warburg-like effect in Caenorhabditis elegans. Toxicol. Sci. 2016, 152, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Ajima, M.N.; Pandey, P.K.; Kumar, K.; Poojary, N. Neurotoxic effects, molecular responses and oxidative stress biomarkers in Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) exposed to verapamil. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 196, 44–52. [Google Scholar] [CrossRef]
- El Hajam, M.; Plavan, G.I.; Kandri, N.I.; Dumitru, G.; Nicoara, M.N.; Zerouale, A.; Faggio, C. Evaluation of softwood and hardwood sawmill wastes impact on the common carp “Cyprinus carpio” and its aquatic environment: An oxidative stress study. Environ. Toxicol. Pharmacol. 2020, 75, 103327. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, H.; Sung, G.; Seo, S.A.; Kim, K.I.; Kang, Y.J.; Kang, J.C. Toxic effects on hematological parameters and oxidative stress in juvenile olive flounder, Paralichthys olivaceus exposed to waterborne zinc. Aquac. Rep. 2019, 15, 100225. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, L.; Wang, J.; Xie, H.; Wang, J.; Han, Y.; Yang, J. Oxidative stress and lipid peroxidation in the earthworm Eisenia fetida induced by low doses of fomesafen. Environ. Sci. Pollut. Res. 2013, 20, 201–208. [Google Scholar] [CrossRef]
- Widiastuti, I.M.; Hertika, A.M.S.; Musa, M.; Arfiati, D. Superdioxide dismutase (SOD) and metallothionein (MT) Tubifex tubifex at the acute mercury exposure. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 236, p. 012101. [Google Scholar]
- Stevens, F.C. Calmodulin: An introduction. Can. J. Biochem. Cell Biol. = Rev. Can. Biochim. Biol. Cell. 1983, 61, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, F.; Liao, Y.; Jin, Y.; Sun, G. Effects of arsenite in astrocytes on neuronal signaling transduction. Toxicology 2013, 303, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, X.; Liu, X.; Jiang, L.; Yang, G.; Shi, X.; Zhang, C.; Piao, F. Inhibition of miR-219 alleviates arsenic-induced learning and memory impairments and synaptic damage through up-regulating CaMKII in the hippocampus. Neurochem. Res. 2018, 43, 948–958. [Google Scholar] [CrossRef]
- Robison, A.J.; Winder, D.G.; Colbran, R.J.; Bartlett, R.K. Oxidation of calmodulin alters activation and regulation of CaMKII. Biochem. Biophys. Res. Commun. 2007, 356, 97–101. [Google Scholar] [CrossRef]
- Pang, X.; Zhang, P.; Chen, X.; Liu, W. Ubiquitin-proteasome pathway in skeletal muscle atrophy. Front. Physiol. 2023, 14, 1289537. [Google Scholar] [CrossRef]
- Della Sala, G.; Agriesti, F.; Mazzoccoli, C.; Tataranni, T.; Costantino, V.; Piccoli, C. Clogging the Ubiquitin-Proteasome Machinery with Marine Natural Products: Last Decade Update. Mar. Drugs 2018, 16, 467. [Google Scholar] [CrossRef]
- Ardley, H.C.; Scott, G.B.; Rose, S.A.; Tan, N.G.; Markham, A.F.; Robinson, P.A. Inhibition of proteasomal activity causes inclusion formation in neuronal and non-neuronal cells overexpressing Parkin. Mol. Biol. Cell 2003, 14, 4541–4556. [Google Scholar] [CrossRef]
- Planelló, R.; Martínez-Guitarte, J.L.; Morcillo, G. Ribosomal genes as early targets of cadmium-induced toxicity in Chironomus riparius larvae. Sci. Total Environ. 2007, 373, 113–121. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Choi, J. Characterization of a ribosomal protein L15 cDNA from Chironomus riparius (Diptera; Chironomidae): Transcriptional regulation by cadmium and silver nanoparticles. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 159, 157–162. [Google Scholar] [CrossRef]
- Herrero, Ó.; Morcillo, G.; Planelló, R. Transcriptional deregulation of genetic biomarkers in Chironomus riparius larvae exposed to ecologically relevant concentrations of di (2-ethylhexyl) phthalate (DEHP). PLoS ONE 2017, 12, e0171719. [Google Scholar] [CrossRef]
- Méndez-Fernández, L.; Martínez-Madrid, M.; Rodríguez, P. Toxicity and critical body residues of Cd, Cu and Cr in the aquatic oligochaete Tubifex tubifex (Müller) based on lethal and sublethal effects. Ecotoxicology 2013, 22, 1445–1460. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Moogoui, R.; Gupta, D.K. Arsenic: Source, Occurrence, Cycle, and Detection. In Arsenic Contamination in the Environment: The Issues and Solutions; Gupta, D.K., Chatterjee, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 13–35. [Google Scholar]
- Chapman, P.M.; Farrell, M.A.; Brinkhurst, R.O. Relative tolerances of selected aquatic oligochaetes to individual pollutants and environmental factors. Aquat. Toxicol. 1982, 2, 47–67. [Google Scholar] [CrossRef]
- APHA, American Public Health Association. Part 8510. Toxicity test procedures for annelids. In Standard Methods for the Examination of Water and Wastewaters, 17th ed.; American Public Health Association (APHA): Washington, DC, USA, 1989. [Google Scholar]
- Clements, W.H.; Kiffney, P.M. Integrated laboratory and field approach for assessing impacts of heavy metals at the Arkansas River, Colorado. Environ. Toxicol. Chem. Int. J. 1994, 13, 397–404. [Google Scholar] [CrossRef]
- NIST 2976; Standard Reference Material 2976. National Institute of Standards & Technology: Gaithersburg, MD, USA, 2008.
- TMDA 52.3; Certified Reference Materials (CRMs). Environment Canada National Laboratory for Environmental Testing: Burlington, ON, Canada, 2010.
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 5 March 2024).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization, and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, R.; Barrett, T.; Beck, J.; Benson, D.A.; Bollin, C.; Bolton, E.; Bourexis, D.; Brister, J.R.; Bryant, S.H.; Canese, K.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- IBM® SPSS Statistics. SPSS 24.0 for Windows, SPSS Inc. IBM Company: Armonk, NY, USA, 2016.
- Hamilton, M.A.; Russo, R.C.; Thurston, R.V. Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ. Sci. Technol. 1977, 11, 714–719. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]


| As (mg L−1) | Mortality | Autotomy | ||||||
|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| 0.00 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| 38 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 63 | 0 | 0 | 0 | 0 | 2 | 2 | 2 | 2 |
| 100 | 0 | 2 | 6 | 11 | 2 | 8 | 12 | 12 |
| 159 | 4 | 7 | 11 | 13 | 6 | 11 | 13 | 15 |
| 250 | 2 | 9 | 18 | 18 | 8 | 18 | 18 | 18 |
| 407 | 12 | 14 | 15 | 16 | 13 | 15 | 16 | 16 |
| Mortality | Autotomy | |||||||
|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| NOEC | 250 | - | 159 | 159 | - | 159 | 159 | 159 |
| LOEC | 407 | ns | 250 | 250 | ns | 250 | 250 | 250 |
| (p value) | (0.041) | - | (0.019) | (0.028) | - | (0.013) | (0.011) | (0.015) |
| LC50/EC50 | >407 | 260 | 163 | 137 * | 387 | 149 | 126 | 120 |
| 95% CI | - | 210–349 | 135–199 | 136–137 | 280–718 | 120–188 | 101–155 | 96–148 |
| Mortality | Autotomy | |||||||
|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| LR50/ER50 | 1072 | - | 679 * | 578 * | 1019 | 634 * | 555 * | 479 * |
| 95% CI | 891–1502 | - | 671–687 | 574–582 | 795–1623 | 630–638 | 553–557 | 477–481 |
| Gene | Primer Sequence (5′–3′) | Fragment Size (bp) |
|---|---|---|
| Hsc70 | F ATGGACAAGCCGGCCATCCACG R GAGGGACAGCGGAGCAACGT | 228 |
| Hsp10 | F AAAGGTGCTCGAAGCGACAG R ACGACTGGAATTTGCCGAGA | 191 |
| Hsp60 | F GGTCCAAGACATCGCACACA R GGCTTCCATCACACCTCGTC | 151 |
| Hsp83 | F GAGCAGATGGAGGACGGAGA R CGAATCTTGTCGAGGGCATC | 152 |
| COX1 | F CGGGTGTATGCTTAGCAAACTCA R CCGAATACTGCCCCCATTCT | 103 |
| Cat | F GTGCTGAACCGTAGCCCAAA R ACGAGAACAGACGACCCTGAAG | 124 |
| GSR | F ACCGTTGTGTTCAGCCATCC R TTCTTTTGTGTCATTGCGTGGT | 137 |
| MnSOD | F TGCCGAAGCACAGGCTAAA R CTCAAGCGAACCAAAGTCACG | 184 |
| CaM | F AAGGAACTGGGGACCGTGAT R TCAGGAACTCGGGAAAGTCTATTG | 121 |
| RpS13 | F TTGGCGTTATTCTGCGTGACT R TTGTCCTTGCGGTTCCTCTC | 178 |
| UBE2 | F CGTCTGCTTGTGGTGGTGAC R TCGTTGTCCATCTCGTCGTAATAAT | 118 |
| Gapdh | F GGCAAGCTGACTGGAATGGC R TGCCGCTTTGATTTGGTCGT | 108 |
| 28S | F TTCGCGACCTCAACTCATGT R CCGCATTCAAGCTGGACTTA | 220 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Ocio, I.; Aquilino, M.; Llorente, L.; Martínez-Madrid, M.; Rodríguez, P.; Méndez-Fernández, L.; Planelló, R. Toxicogenomics of the Freshwater Oligochaete, Tubifex tubifex (Annelida, Clitellata), in Acute Water-Only Exposure to Arsenic. Int. J. Mol. Sci. 2024, 25, 3382. https://doi.org/10.3390/ijms25063382
Moreno-Ocio I, Aquilino M, Llorente L, Martínez-Madrid M, Rodríguez P, Méndez-Fernández L, Planelló R. Toxicogenomics of the Freshwater Oligochaete, Tubifex tubifex (Annelida, Clitellata), in Acute Water-Only Exposure to Arsenic. International Journal of Molecular Sciences. 2024; 25(6):3382. https://doi.org/10.3390/ijms25063382
Chicago/Turabian StyleMoreno-Ocio, Iñigo, Mónica Aquilino, Lola Llorente, Maite Martínez-Madrid, Pilar Rodríguez, Leire Méndez-Fernández, and Rosario Planelló. 2024. "Toxicogenomics of the Freshwater Oligochaete, Tubifex tubifex (Annelida, Clitellata), in Acute Water-Only Exposure to Arsenic" International Journal of Molecular Sciences 25, no. 6: 3382. https://doi.org/10.3390/ijms25063382