
The Contribution of the Zebrafish Model to the Understanding of Polycomb Repression in Vertebrates

Abstract
:1. Introduction
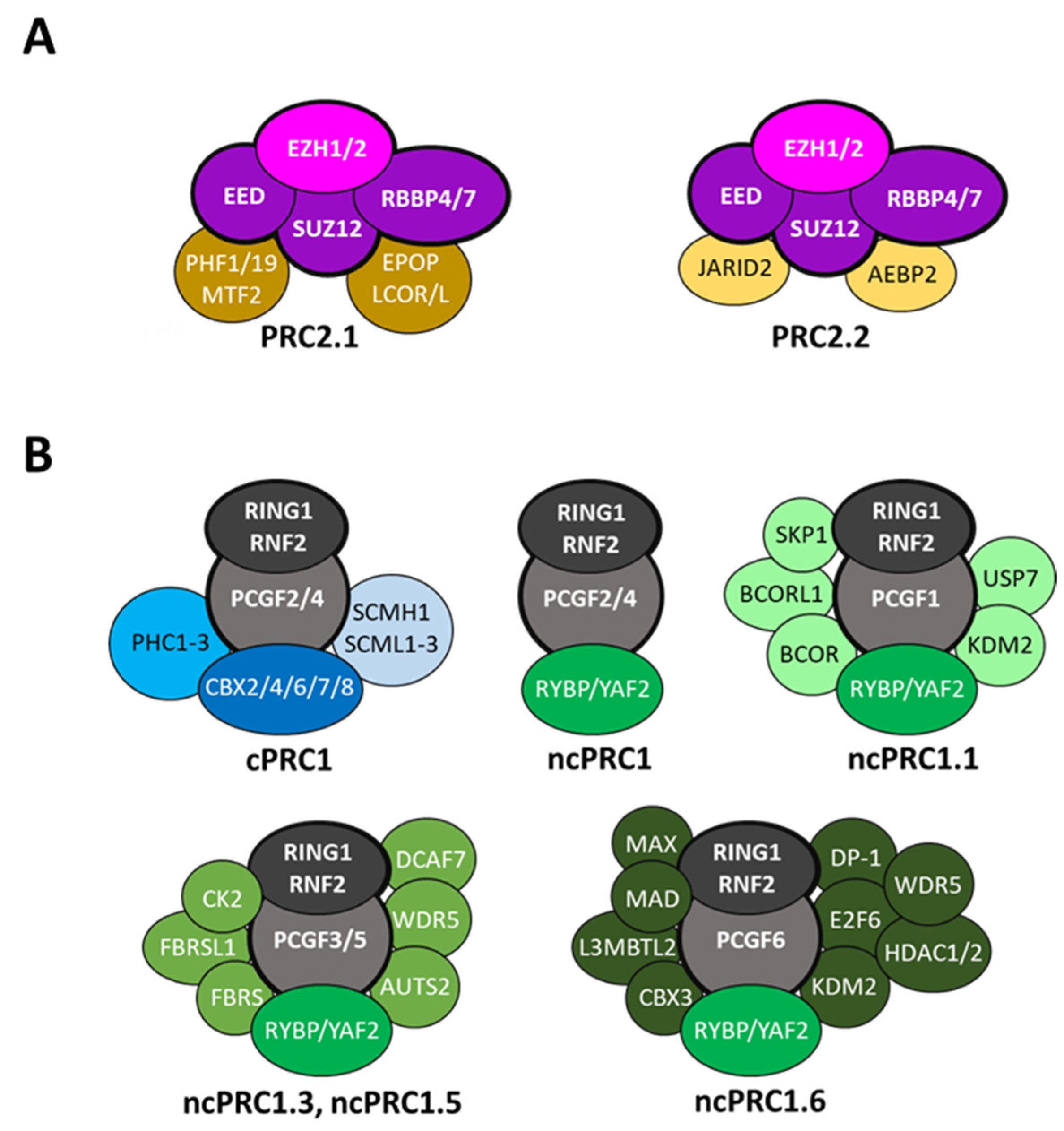
2. The Zebrafish Genes Coding for the Components of the PRC1 and PRC2 Complexes
3. Genetic Approaches to Study Zebrafish Polycomb Group Gene Function
4. Polycomb Group Proteins Support the Development and Viability of Zebrafish
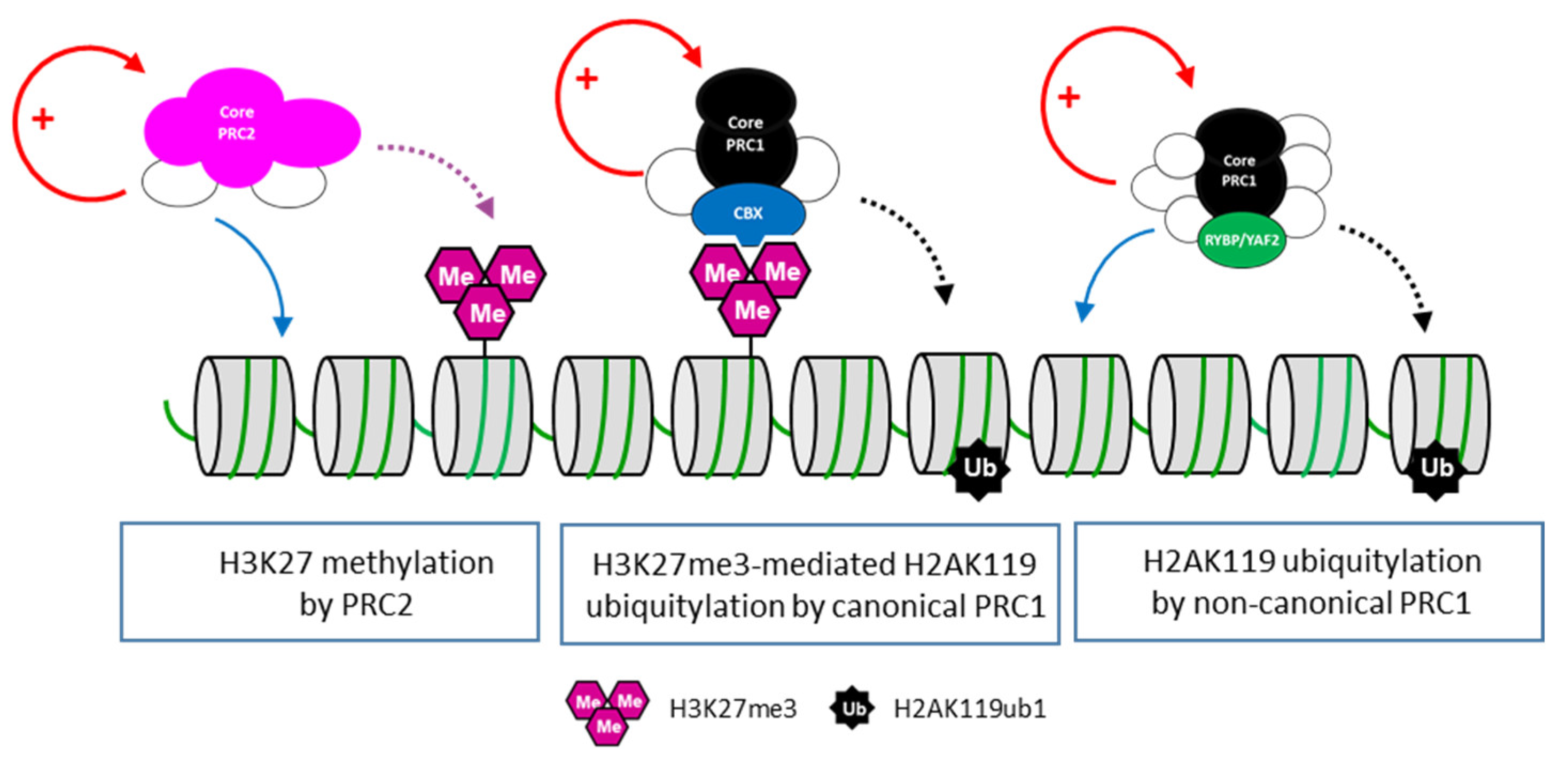
4.1. Role of PRC1 in Zebrafish Development
4.2. Role of PRC2 in Zebrafish Development
4.3. Function of Polycomb Repression in Zebrafish Digestive Tissues
4.4. Polycomb Repression in Cardiac Cell Identity and Function
4.5. Polycomb Repression in Zebrafish Developmental Hematopoiesis
4.6. Polycomb Repression and Nervous System Development in Zebrafish
4.7. Polycomb Repression in Cancer
4.8. Polycomb Repression and Behavior
5. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene | Morphant/Mutant | Phenotype | References |
|---|---|---|---|
| ezh1 | MO-ezh1 | Mild decrease in primitive erythropoiesis | [140] |
| MO-ezh1 | Increase in hematopoietic stem and progenitor cells via enhanced commitment to hemogenic fate | [68] | |
| ezh1(ul3) | Viable, fertile, without obvious phenotype | [69] | |
| ezh1(b1394) | Viable, fertile, without obvious phenotype | [70] | |
| ezh2 | ezh2(hu5670) | Lethal around 9–12 dpf. Intestine and liver defects | [137] |
| MZezh2(hu5670) | Lethal around 2 dpf. Heart, liver and pancreas defects | [72] | |
| ezh2(ul2) | Lethal around 12 dpf. Intestine defects, liver steatosis, smaller pancreas | [73] | |
| ezh2(sal1199) | Hypormophic allele. Viable with an open mouth phenotype (prominent lower jaw) and mild appendage patterning defects | [70,74] | |
| ezh2(b1392) | Lethal between 7 and 14 dpf. Craniofacial defects (reduced branchiostegal ray 3 length) | [70] | |
| eed | MO-eed | Intermediate decrease in definitive hematopoietic stem cells | [140] |
| eed(ul4) | Lethal around 10–12 dpf. Reduced size of the pancreas, hepatic steatosis and a loss of intestinal structures. Neuronal differentiation defects. Hyperactive behavor | [75] | |
| suz12a/b | MO-suz12b | Intermediate decrease in primitive erythropoiesis and definitive hematopoietic stem cells | [140] |
| suz12a;suz12b | Lethal between 8 and 15 dpf (suz12a-/-; suz12b-/-) | [76] | |
| rbbp4 | MO-rbbp4 | Intermediate decrease in primitive erythropoiesis | [140] |
| MO-rbbp4 | Impaired fin regeneration | [77] | |
| rbbp4(is60) | Lethal between 5 and 10 dpf. Microcephaly and microphtalmia | [78] | |
| skp1 | skp1(hi3970Tg) | Small head and eyes, underdeveloped liver/gut | [93] |
| jarid2a/b | MO-jarid2a | Intermediate decrease in primitive erythropoiesis | [140] |
| MO-jarid2b | Intermediate decrease in primitive erythropoiesis | [140] | |
| rnf2 | MO-rnf2 | Reduction in definitive hematopoietic stem cells | [79] |
| rnf2(ibl31) | Lethal at around 4–5 dpf. Absence of pectoral fins, craniofacial defects, smaller intestine, absence of terminal intestinal differentiation in the liver, absence of pancreatic lobe, heart defects | [80,111,139] | |
| rnf2(f5)–rnf2(f8) | Defective craniofacial structures, small eyes, absence of pectoral fins, pericardial edema. Disruption of cardiac sarcomere assembly. Abnormal development of central and enteric nervous systems | [81,141] | |
| pcgf1 | MO-pcgf1 | Small head, reduced (or even absent) telencephalon, abnormal activation of the neural induction process | [82] |
| pcgf1(ul1) | Viable, fertile, early signs of aging | [83] | |
| bmi1a/b | MO-bmi1a/b | Microcephaly | [85] |
| MO-bmi1a/b | Reduction in definitive hematopoietic stem cells | [79] | |
| cbx2 | MO-cbx2 | Intermediate decrease in primitive erythropoiesis | [140] |
| cbx6a/b | MO-cbx6a | Mild decrease in primitive erythropoiesis and intermediate decrease in definitive hematopoietic stem cells | [140] |
| MO-cbx6b | Strong decrease in definitive hematopoietic stem cells | [140] | |
| cbx7a | MO-cbx7a | Mild decrease in primitive erythropoiesis | [140] |
| cbx8a/b | MO-cbx8a | Intermediate decrease in primitive erythropoiesis | [140] |
| MO-cbx8b | Intermediate decrease in primitive erythropoiesis and strong decrease in definitive hematopoietic stem cells | [140] | |
| phc2a | MO-phc2a | Abnormal epibolic movements and a thick tailbud or incomplete covering of the yolk plug | [87] |
| yaf2 | MO-yaf2 | Developmental arrest before the one-somite stage. Central nervous system degeneration | [88] |
| bcor | MO-bcor | Medial displacement of the eyes and severe colobomata. Abnormal neural tube development | [89,90] |
| kdm2bb | MO-kdm2bb | Early embryonic lethality at 24 hpf at high MO doses, and no phenotype at low MO doses | [91] |
| usp7 | MO-usp7 | Alteration in craniofacial development (mild changes on the branchial arches) | [92] |
| wdr5 | MO-wdr5 | Intermediate decrease in primitive erythropoiesis | [140] |
| MO-wdr5 | Small liver phenotype | [94] | |
| wdr5(zju131) | Lethal at around 8 dpf. Curved body and undeveloped liver | [94] | |
| auts2a | auts2a(ncb104) | Viable and normal morphology. High variability and delayed escape behavior | [95] |
| mgaa | MO-mgaa | Severe brain abnormalities, small eyes, failure in heart tube looping with edema, lack of pigmentation, reduced levels of circulating erythrocytes | [96] |
| mgaa(ihb801) | Reduction of ventral tail fin at 1 dpf | [97] |
References
- Kassis, J.A.; Kennison, J.A.; Tamkun, J.W. Polycomb and Trithorax Group Genes in Drosophila. Genetics 2017, 206, 1699–1725. [Google Scholar] [CrossRef] [Green Version]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.S.; Weiss, A.; Erdjument-Bromage, H.; Shao, Z.; Tempst, P.; Kingston, R.E. The core of the polycomb repressive complex is compositionally and functionally conserved in flies and humans. Mol. Cell. Biol. 2002, 22, 6070–6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Bracken, A.P.; Dietrich, N.; Pasini, D.; Hansen, K.H.; Helin, K. Genome-wide mapping of Polycomb target genes unravels their roles in cell fate transitions. Genes Dev. 2006, 20, 1123–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolhuis, B.; de Wit, E.; Muijrers, I.; Teunissen, H.; Talhout, W.; van Steensel, B.; van Lohuizen, M. Genome-wide profiling of PRC1 and PRC2 Polycomb chromatin binding in Drosophila melanogaster. Nat. Genet. 2006, 38, 694–699. [Google Scholar] [CrossRef]
- Varlet, E.; Ovejero, S.; Martinez, A.M.; Cavalli, G.; Moreaux, J. Role of Polycomb Complexes in Normal and Malignant Plasma Cells. Int. J. Mol. Sci. 2020, 21, 8047. [Google Scholar] [CrossRef]
- Piunti, A.; Shilatifard, A. The roles of Polycomb repressive complexes in mammalian development and cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Scholl, A.; De, S. Epigenetic Regulation by Polycomb Complexes from Drosophila to Human and Its Relation to Communicable Disease Pathogenesis. Int. J. Mol. Sci. 2022, 23, 12285. [Google Scholar] [CrossRef]
- Bölicke, N.; Albert, M. Polycomb-mediated gene regulation in human brain development and neurodevelopmental disorders. Dev. Neurobiol. 2022, 82, 345–363. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.J.; Xu, J.L.; Qi, Y.R.; Yuan, Z.Q.; Zhao, W. Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer. Int. J. Mol. Sci. 2022, 23, 9574. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, L.; Erdjument-Bromage, H.; Vidal, M.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of histone H2A ubiquitination in Polycomb silencing. Nature 2004, 431, 873–878. [Google Scholar] [CrossRef]
- de Napoles, M.; Mermoud, J.E.; Wakao, R.; Tang, Y.A.; Endoh, M.; Appanah, R.; Nesterova, T.B.; Silva, J.; Otte, A.P.; Vidal, M.; et al. Polycomb group proteins Ring1A/B link ubiquitylation of histone H2A to heritable gene silencing and X inactivation. Dev. Cell 2004, 7, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Wang, L.; Wang, H.; Xia, L.; Erdjument-Bromage, H.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 2002, 298, 1039–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmichev, A.; Nishioka, K.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Histone methyltransferase activity associated with a human multiprotein complex containing the Enhancer of Zeste protein. Genes Dev. 2002, 16, 2893–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.; Hart, C.M.; Francis, N.J.; Vargas, M.L.; Sengupta, A.; Wild, B.; Miller, E.L.; O’Connor, M.B.; Kingston, R.E.; Simon, J.A. Histone methyltransferase activity of a Drosophila Polycomb group repressor complex. Cell 2002, 111, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, Y.B.; Kahn, T.G.; Nix, D.A.; Li, X.Y.; Bourgon, R.; Biggin, M.; Pirrotta, V. Genome-wide analysis of Polycomb targets in Drosophila melanogaster. Nat. Genet. 2006, 38, 700–705. [Google Scholar] [CrossRef]
- Fursova, N.A.; Blackledge, N.P.; Nakayama, M.; Ito, S.; Koseki, Y.; Farcas, A.M.; King, H.W.; Koseki, H.; Klose, R.J. Synergy between Variant PRC1 Complexes Defines Polycomb-Mediated Gene Repression. Mol. Cell 2019, 74, 1020–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuettengruber, B.; Bourbon, H.M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef] [Green Version]
- Blackledge, N.P.; Klose, R.J. The molecular principles of gene regulation by Polycomb repressive complexes. Nat. Rev. Mol. Cell. Biol. 2021, 22, 815–833. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, G.; Veenstra, G.J.C.; Vermeulen, M.; Marks, H. The Complexity of PRC2 Subcomplexes. Trends Cell Biol. 2019, 29, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.R.; Lee, C.H.; Oksuz, O.; Stafford, J.M.; Reinberg, D. PRC2 is high maintenance. Genes Dev. 2019, 33, 903–935. [Google Scholar] [CrossRef] [PubMed]
- Glancy, E.; Ciferri, C.; Bracken, A.P. Structural basis for PRC2 engagement with chromatin. Curr. Opin. Struct. Biol. 2021, 67, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Weber, L.M.; Liefke, R. Evolutionary adaptation of the Polycomb repressive complex 2. Epigenetics Chromatin 2022, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.G. No Easy Way Out for EZH2: Its Pleiotropic, Noncanonical Effects on Gene Regulation and Cellular Function. Int. J. Mol. Sci. 2020, 21, 9501. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, J.; Bonasio, R.; Strino, F.; Sawai, A.; Parisi, F.; Kluger, Y.; Reinberg, D. PCGF homologs, CBX proteins, and RYBP define functionally distinct PRC1 family complexes. Mol. Cell 2012, 45, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Illingworth, R.S. Chromatin folding and nuclear architecture: PRC1 function in 3D. Curr. Opin. Genet. Dev. 2019, 55, 82–90. [Google Scholar] [CrossRef]
- Geng, Z.; Gao, Z. Mammalian PRC1 Complexes: Compositional Complexity and Diverse Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 8594. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Dimitrova, E.; Oxley, D.; Webster, J.; Poot, R.; Demmers, J.; Bezstarosti, K.; Taylor, S.; Ura, H.; Koide, H.; et al. RYBP-PRC1 complexes mediate H2A ubiquitylation at polycomb target sites independently of PRC2 and H3K27me3. Cell 2012, 148, 664–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morey, L.; Aloia, L.; Cozzuto, L.; Benitah, S.A.; Di Croce, L. RYBP and Cbx7 define specific biological functions of polycomb complexes in mouse embryonic stem cells. Cell Rep. 2013, 3, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.R.; King, H.W.; Blackledge, N.P.; Fursova, N.A.; Ember, K.J.; Fischer, R.; Kessler, B.M.; Klose, R.J. RYBP stimulates PRC1 to shape chromatin-based communication between Polycomb repressive complexes. Elife 2016, 5, e18591. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, N.P.; Zhou, J.C.; Tolstorukov, M.Y.; Farcas, A.M.; Park, P.J.; Klose, R.J. CpG islands recruit a histone H3 lysine 36 demethylase. Mol. Cell 2010, 38, 179–190. [Google Scholar] [CrossRef]
- He, J.; Shen, L.; Wan, M.; Taranova, O.; Wu, H.; Zhang, Y. Kdm2b maintains murine embryonic stem cell status by recruiting PRC1 complex to CpG islands of developmental genes. Nat. Cell Biol. 2013, 15, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farcas, A.M.; Blackledge, N.P.; Sudbery, I.; Long, H.K.; McGouran, J.F.; Rose, N.R.; Lee, S.; Sims, D.; Cerase, A.; Sheahan, T.W.; et al. KDM2B links the Polycomb Repressive Complex 1 (PRC1) to recognition of CpG islands. Elife 2012, 1, e00205. [Google Scholar] [CrossRef]
- Jolma, A.; Yan, J.; Whitington, T.; Toivonen, J.; Nitta, K.R.; Rastas, P.; Morgunova, E.; Enge, M.; Taipale, M.; Wei, G.; et al. DNA-binding specificities of human transcription factors. Cell 2013, 152, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaubatz, S.; Wood, J.G.; Livingston, D.M. Unusual proliferation arrest and transcriptional control properties of a newly discovered E2F family member, E2F-6. Proc. Natl. Acad. Sci. USA 1998, 95, 9190–9195. [Google Scholar] [CrossRef] [Green Version]
- Stielow, B.; Finkernagel, F.; Stiewe, T.; Nist, A.; Suske, G. MGA, L3MBTL2 and E2F6 determine genomic binding of the non-canonical Polycomb repressive complex PRC1.6. PLoS Genet. 2018, 14, e1007193. [Google Scholar] [CrossRef] [PubMed]
- Engeszer, R.E.; Patterson, L.B.; Rao, A.A.; Parichy, D.M. Zebrafish in the wild: A review of natural history and new notes from the field. Zebrafish 2007, 4, 21–40. [Google Scholar] [CrossRef]
- Laale, H.W. The biology and use of zebrafish, Brachydanio rerio in fisheries research. A literature review. J. Fish Biol. 1977, 10, 121–173. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Kettleborough, R.N.; Busch-Nentwich, E.M.; Harvey, S.A.; Dooley, C.M.; de Bruijn, E.; van Eeden, F.; Sealy, I.; White, R.J.; Herd, C.; Nijman, I.J.; et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature 2013, 496, 494–497. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K. Transposon tools and methods in zebrafish. Dev. Dyn. 2005, 234, 244–254. [Google Scholar] [CrossRef]
- Bedell, V.M.; Westcot, S.E.; Ekker, S.C. Lessons from morpholino-based screening in zebrafish. Brief. Funct. Genom. 2011, 10, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jia, Z.; Zhang, S.; He, X. Progress in Gene-Editing Technology of Zebrafish. Biomolecules 2021, 11, 1300. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.S.; Van de Peer, Y.; Braasch, I.; Meyer, A. Comparative genomics provides evidence for an ancient genome duplication event in fish. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1661–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoegg, S.; Brinkmann, H.; Taylor, J.S.; Meyer, A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J. Mol. Evol. 2004, 59, 190–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sémon, M.; Wolfe, K.H. Reciprocal gene loss between Tetraodon and zebrafish after whole genome duplication in their ancestor. Trends Genet. 2007, 23, 108–112. [Google Scholar] [CrossRef]
- Brunet, F.G.; Roest Crollius, H.; Paris, M.; Aury, J.-M.; Gibert, P.; Jaillon, O.; Laudet, V.; Robinson-Rechavi, M. Gene loss and evolutionary rates following whole-genome duplication in teleost fishes. Mol. Biol. Evol. 2006, 23, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Le Faou, P.; Völkel, P.; Angrand, P.O. The zebrafish genes encoding the Polycomb repressive complex (PRC) 1. Gene 2011, 475, 10–21. [Google Scholar] [CrossRef]
- Raby, L.; Völkel, P.; Le Bourhis, X.; Angrand, P.O. The Polycomb Orthologues in Teleost Fishes and Their Expression in the Zebrafish Model. Genes 2020, 11, 362. [Google Scholar] [CrossRef]
- Vizán, P.; Beringer, M.; Ballaré, C.; Di Croce, L. Role of PRC2-associated factors in stem cells and disease. FEBS J. 2015, 282, 1723–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Dang, Y.; Hu, B.; Luo, L.; Zhao, P.; Wang, S.; Zhang, K. Overlapping functions of RBBP4 and RBBP7 in regulating cell proliferation and histone H3.3 deposition during mouse preimplantation development. Epigenetics 2022, 17, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhu, Y.; Xing, F.; Liu, S.; Xia, Y.; Jiang, Q.; Qin, J. The polycomb group protein PCGF6 mediates germline gene silencing by recruiting histone-modifying proteins to target gene promoters. J. Biol. Chem. 2020, 295, 9712–9724. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.L.; Hsieh, J.; Barbosa, A.C.; Richardson, J.A.; Olson, E.N. Histone deacetylases 1 and 2 control the progression of neural precursors to neurons during brain development. Proc. Natl. Acad. Sci. USA 2009, 106, 7876–7881. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.L.; Davis, C.A.; Potthoff, M.J.; Haberland, M.; Fielitz, J.; Qi, X.; Hill, J.A.; Richardson, J.A.; Olson, E.N. Histone deacetylases 1 and 2 redundantly regulate cardiac morphogenesis, growth, and contractility. Genes Dev. 2007, 21, 1790–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovey, O.M.; Foster, C.T.; Cowley, S.M. Histone deacetylase 1 (HDAC1), but not HDAC2, controls embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 8242–8247. [Google Scholar] [CrossRef] [Green Version]
- Summerton, J.; Weller, D. Morpholino antisense oligomers: Design, preparation, and properties. Antisense Nucleic Acid Drug Dev. 1997, 7, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bill, B.R.; Petzold, A.M.; Clark, K.J.; Schimmenti, L.A.; Ekker, S.C. A primer for morpholino use in zebrafish. Zebrafish 2009, 6, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robu, M.E.; Larson, J.D.; Nasevicius, A.; Beiraghi, S.; Brenner, C.; Farber, S.A.; Ekker, S.C. p53 activation by knockdown technologies. PLoS Genet. 2007, 3, e78. [Google Scholar] [CrossRef] [Green Version]
- Stainier, D.Y.R.; Kontarakis, Z.; Rossi, A. Making sense of anti-sense data. Dev. Cell 2015, 32, 7–8. [Google Scholar] [CrossRef]
- Kok, F.O.; Shin, M.; Ni, C.W.; Gupta, A.; Grosse, A.S.; van Impel, A.; Kirchmaier, B.C.; Peterson-Maduro, J.; Kourkoulis, G.; Male, I.; et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Dev. Cell 2015, 32, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morcos, P.A.; Vincent, A.C.; Moulton, J.D. Gene Editing Versus Morphants. Zebrafish 2015, 12, 319. [Google Scholar] [CrossRef]
- Zimmer, A.M.; Pan, Y.K.; Chandrapalan, T.; Kwong, R.W.M.; Perry, S.F. Loss-of-function approaches in comparative physiology: Is there a future for knockdown experiments in the era of genome editing? J. Exp. Biol. 2019, 222, jeb175737. [Google Scholar] [CrossRef] [Green Version]
- Salanga, C.M.; Salanga, M.C. Genotype to Phenotype: CRISPR Gene Editing Reveals Genetic Compensation as a Mechanism for Phenotypic Disjunction of Morphants and Mutants. Int. J. Mol. Sci. 2021, 22, 3472. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E.; Schulte-Merker, S.; Walderich, B.; Plasterk, R.H. Target-selected inactivation of the zebrafish rag1 gene. Science 2002, 297, 99–102. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Parant, J.M.; Yehn, J.R. Approaches to Inactivate Genes in Zebrafish. Adv. Exp. Med. Biol. 2016, 916, 61–86. [Google Scholar] [CrossRef]
- Soto, R.A.; Najia, M.A.T.; Hachimi, M.; Frame, J.M.; Yette, G.A.; Lummertz da Rocha, E.; Stankunas, K.; Daley, G.Q.; North, T.E. Sequential regulation of hemogenic fate and hematopoietic stem and progenitor cell formation from arterial endothelium by Ezh1/2. Stem Cell Rep. 2021, 16, 1718–1734. [Google Scholar] [CrossRef]
- Völkel, P.; Bary, A.; Raby, L.; Chapart, A.; Dupret, B.; Le Bourhis, X.; Angrand, P.O. Ezh1 arises from Ezh2 gene duplication but its function is not required for zebrafish development. Sci. Rep. 2019, 9, 4319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yette, G.A.; Stewart, S.; Stankunas, K. Zebrafish Polycomb repressive complex-2 critical roles are largely Ezh2- over Ezh1-driven and concentrate during early embryogenesis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Zhong, Y.; Ye, Q.; Chen, C.; Wang, M.; Wang, H. Ezh2 promotes clock function and hematopoiesis independent of histone methyltransferase activity in zebrafish. Nucleic Acids Res. 2018, 46, 3382–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San, B.; Chrispijn, N.D.; Wittkopp, N.; van Heeringen, S.J.; Lagendijk, A.K.; Aben, M.; Bakkers, J.; Ketting, R.F.; Kamminga, L.M. Normal formation of a vertebrate body plan and loss of tissue maintenance in the absence of ezh2. Sci. Rep. 2016, 6, 24658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupret, B.; Völkel, P.; Vennin, C.; Toillon, R.A.; Le Bourhis, X.; Angrand, P.O. The histone lysine methyltransferase Ezh2 is required for maintenance of the intestine integrity and for caudal fin regeneration in zebrafish. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 1079–1093. [Google Scholar] [CrossRef]
- San, B.; Rougeot, J.; Voeltzke, K.; van Vegchel, G.; Aben, M.; Andralojc, K.M.; Flik, G.; Kamminga, L.M. The ezh2(sa1199) mutant zebrafish display no distinct phenotype. PLoS ONE 2019, 14, e0210217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raby, L.; Völkel, P.; Hasanpour, S.; Cicero, J.; Toillon, R.A.; Adriaenssens, E.; Van Seuningen, I.; Le Bourhis, X.; Angrand, P.O. Loss of Polycomb Repressive Complex 2 Function Alters Digestive Organ Homeostasis and Neuronal Differentiation in Zebrafish. Cells 2021, 10, 3142. [Google Scholar] [CrossRef] [PubMed]
- Oppel, F.; Ki, D.H.; Zimmerman, M.W.; Ross, K.N.; Tao, T.; Shi, H.; He, S.; Aster, J.C.; Look, A.T. suz12 inactivation in p53- and nf1-deficient zebrafish accelerates the onset of malignant peripheral nerve sheath tumors and expands the spectrum of tumor types. Dis. Model Mech. 2020, 13, dmm042341. [Google Scholar] [CrossRef]
- Pfefferli, C.; Müller, F.; Jaźwińska, A.; Wicky, C. Specific NuRD components are required for fin regeneration in zebrafish. BMC Biol. 2014, 12, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, L.E.; Haltom, J.A.; Almeida, M.P.; Wierson, W.A.; Solin, S.L.; Weiss, T.J.; Helmer, J.A.; Sandquist, E.J.; Shive, H.R.; McGrail, M. Epigenetic regulators Rbbp4 and Hdac1 are overexpressed in a zebrafish model of RB1 embryonal brain tumor, and are required for neural progenitor survival and proliferation. Dis. Model Mech. 2018, 11, dmm034124. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Mazor, T.; Huang, H.; Huang, H.T.; Kathrein, K.L.; Woo, A.J.; Chouinard, C.R.; Labadorf, A.; Akie, T.E.; Moran, T.B.; et al. Direct recruitment of polycomb repressive complex 1 to chromatin by core binding transcription factors. Mol. Cell 2012, 45, 330–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Velden, Y.U.; Wang, L.; van Lohuizen, M.; Haramis, A.P. The Polycomb group protein Ring1b is essential for pectoral fin development. Development 2012, 139, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Feng, G.; Zhang, Y.; Sun, Y. PRC1 Stabilizes Cardiac Contraction by Regulating Cardiac Sarcomere Assembly and Cardiac Conduction System Construction. Int. J. Mol. Sci. 2021, 22, 11368. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ji, G.; Zhou, J.; Du, J.; Li, X.; Shi, W.; Hu, Y.; Zhou, W.; Hao, A. Pcgf1 Regulates Early Neural Tube Development Through Histone Methylation in Zebrafish. Front. Cell Dev. Biol. 2021, 8, 581636. [Google Scholar] [CrossRef] [PubMed]
- Dupret, B.; Völkel, P.; Le Bourhis, X.; Angrand, P.O. The Polycomb Group Protein Pcgf1 Is Dispensable in Zebrafish but Involved in Early Growth and Aging. PLoS ONE 2016, 11, e0158700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Wang, L.; Zhu, K.Y.; Dong, M.; Xu, P.F.; Chen, Y.; Chen, S.J.; Chen, Z.; Deng, M.; Liu, T.X. Dominant-negative C/ebpα and polycomb group protein Bmi1 extend short-lived hematopoietic stem/progenitor cell life span and induce lethal dyserythropoiesis. Blood 2011, 118, 3842–3852. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.K.; Wiskow, O.; King, M.J.; Ismail, N.; Ergun, A.; Wang, Y.; Plys, A.J.; Davis, C.P.; Kathrein, K.; Sadreyev, R.; et al. CAT7 and cat7l Long Non-coding RNAs Tune Polycomb Repressive Complex 1 Function during Human and Zebrafish Development. J. Biol. Chem. 2016, 291, 19558–19572. [Google Scholar] [CrossRef] [Green Version]
- Espanola, S.G.; Song, H.; Ryu, E.; Saxena, A.; Kim, E.S.; Manegold, J.E.; Nasamran, C.A.; Sahoo, D.; Oh, C.K.; Bickers, C.; et al. Haematopoietic stem cell-dependent Notch transcription is mediated by p53 through the Histone chaperone Supt16h. Nat. Cell Biol. 2020, 22, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Komoike, Y.; Kawamura, A.; Shindo, N.; Sato, C.; Satoh, J.; Shiurba, R.; Higashinakagawa, T. Zebrafish Polycomb group gene ph2alpha is required for epiboly and tailbud formation acting downstream of FGF signaling. Biochem. Biophys. Res. Commun. 2005, 328, 858–866. [Google Scholar] [CrossRef]
- Stanton, S.E.; McReynolds, L.J.; Evans, T.; Schreiber-Agus, N. Yaf2 inhibits caspase 8-mediated apoptosis and regulates cell survival during zebrafish embryogenesis. J. Biol. Chem. 2006, 281, 28782–28793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Lee, B.K.; Gross, J.M. Bcl6a function is required during optic cup formation to prevent p53-dependent apoptosis and colobomata. Hum. Mol. Genet. 2013, 22, 3568–3582. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.; Thakker, N.; Corcoran, C.M.; Donnai, D.; Perveen, R.; Schneider, A.; Hadley, D.W.; Tifft, C.; Zhang, L.; Wilkie, A.O.; et al. Oculofaciocardiodental and Lenz microphthalmia syndromes result from distinct classes of mutations in BCOR. Nat. Genet. 2004, 36, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Balow, S.A.; Pierce, L.X.; Zentner, G.E.; Conrad, P.A.; Davis, S.; Sabaawy, H.E.; McDermott, B.M., Jr.; Scacheri, P.C. Knockdown of fbxl10/kdm2bb rescues chd7 morphant phenotype in a zebrafish model of CHARGE syndrome. Dev. Biol. 2013, 382, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tse, W.K.F. Importance of deubiquitinases in zebrafish craniofacial development. Biochem. Biophys. Res. Commun. 2017, 487, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Nissen, R.M.; Sun, Z.; Swindell, E.C.; Farrington, S.; Hopkins, N. Identification of 315 genes essential for early zebrafish development. Proc. Natl. Acad. Sci. USA 2004, 101, 12792–12797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Zhu, P.; Shi, H.; Guo, L.; Zhang, Q.; Chen, Y.; Chen, S.; Zhang, Z.; Peng, J.; Chen, J. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature 2019, 568, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.; Kondrychyn, I.; Korzh, V.; Thirumalai, V. High Behavioral Variability Mediated by Altered Neuronal Excitability in auts2 Mutant Zebrafish. eNeuro 2021, 8, ENEURO.0493–20.2021. [Google Scholar] [CrossRef] [PubMed]
- Rikin, A.; Evans, T. The tbx/bHLH transcription factor mga regulates gata4 and organogenesis. Dev. Dyn. 2010, 239, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Chen, J.; Zhang, Y.; Munisha, M.; Dougan, S.; Sun, Y. Mga Modulates Bmpr1a Activity by Antagonizing Bs69 in Zebrafish. Front. Cell Dev. Biol. 2018, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Y.; Harris, M.P.; Levesque, M.P.; Nüsslein-Volhard, C.; Sonawane, M. Heterogeneity across the dorso-ventral axis in zebrafish EVL is regulated by a novel module consisting of sox, snail1a and max genes. Mech. Dev. 2012, 129, 13–23. [Google Scholar] [CrossRef]
- Cunliffe, V.T. Histone deacetylase 1 is required to repress Notch target gene expression during zebrafish neurogenesis and to maintain the production of motoneurones in response to hedgehog signalling. Development 2004, 131, 2983–2995. [Google Scholar] [CrossRef] [Green Version]
- Amsterdam, A.; Burgess, S.; Golling, G.; Chen, W.; Sun, Z.; Townsend, K.; Farrington, S.; Haldi, M.; Hopkins, N. A large-scale insertional mutagenesis screen in zebrafish. Genes Dev. 1999, 13, 2713–2724. [Google Scholar] [CrossRef]
- McCabe, M.T.; Ott, H.M.; Ganji, G.; Korenchuk, S.; Thompson, C.; Van Aller, G.S.; Liu, Y.; Graves, A.P.; Della Pietra, A., 3rd; Diaz, E.; et al. EZH2 inhibition as a therapeutic strategy for lymphoma with EZH2-activating mutations. Nature 2012, 492, 108–112. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Selvaraju, S.; Curtin, M.L.; Jakob, C.G.; Zhu, H.; Comess, K.M.; Shaw, B.; The, J.; Lima-Fernandes, E.; Szewczyk, M.M.; et al. The EED protein-protein interaction inhibitor A-395 inactivates the PRC2 complex. Nat. Chem. Biol. 2017, 13, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Zhao, K.; Gu, J.; Huang, Y.; Wang, Y.; Zhang, H.; Zhang, M.; Zhang, J.; Yu, Z.; Li, L.; et al. An allosteric PRC2 inhibitor targeting the H3K27me3 binding pocket of EED. Nat. Chem. Biol. 2017, 13, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Bird, G.H.; Neff, T.; Guo, G.; Kerenyi, M.A.; Walensky, L.D.; Orkin, S.H. Targeted disruption of the EZH2-EED complex inhibits EZH2-dependent cancer. Nat. Chem. Biol. 2013, 9, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Chen, L.; Jiao, L.; Jiang, X.; Lian, F.; Lu, J.; Zhu, K.; Du, D.; Liu, J.; Ding, H.; et al. Astemizole arrests the proliferation of cancer cells by disrupting the EZH2-EED interaction of polycomb repressive complex 2. J. Med. Chem. 2014, 57, 9512–9521. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gao, S.; Li, J.; Liu, D.; Sheng, C.; Yao, C.; Jiang, W.; Wu, J.; Chen, S.; Huang, W. Wedelolactone disrupts the interaction of EZH2-EED complex and inhibits PRC2-dependent cancer. Oncotarget 2015, 6, 13049–13059. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Ying, W.; Gray, F.; Yao, Y.; Simes, M.L.; Zhao, Q.; Miao, H.; Cho, H.J.; González-Alonso, P.; Winkler, A.; et al. Small-molecule inhibitors targeting Polycomb repressive complex 1 RING domain. Nat. Chem. Biol. 2021, 17, 784–793. [Google Scholar] [CrossRef]
- den Broeder, M.J.; Ballangby, J.; Kamminga, L.M.; Aleström, P.; Legler, J.; Lindeman, L.C.; Kamstra, J.H. Inhibition of methyltransferase activity of enhancer of zeste 2 leads to enhanced lipid accumulation and altered chromatin status in zebrafish. Epigenetics Chromatin 2020, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Mar Lorente, M.; Marcos-Gutiérrez, C.; Pérez, C.; Schoorlemmer, J.; Ramírez, A.; Magin, T.; Vidal, M. Loss- and gain-of-function mutations show a polycomb group function for Ring1A in mice. Development 2000, 127, 5093–5100. [Google Scholar] [CrossRef]
- Voncken, J.W.; Roelen, B.A.; Roefs, M.; de Vries, S.; Verhoeven, E.; Marino, S.; Deschamps, J.; van Lohuizen, M. Rnf2 (Ring1b) deficiency causes gastrulation arrest and cell cycle inhibition. Proc. Natl. Acad. Sci. USA 2003, 100, 2468–2473. [Google Scholar] [CrossRef]
- van der Velden, Y.U.; Wang, L.; Querol Cano, L.; Haramis, A.P. The polycomb group protein ring1b/rnf2 is specifically required for craniofacial development. PLoS ONE 2013, 8, e73997. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ma, Y.; Shang, Y.; Huo, R.; Li, W. Post-translational regulation of the maternal-to-zygotic transition. Cell. Mol. Life Sci. 2018, 75, 1707–1722. [Google Scholar] [CrossRef] [PubMed]
- Posfai, E.; Kunzmann, R.; Brochard, V.; Salvaing, J.; Cabuy, E.; Roloff, T.C.; Liu, Z.; Tardat, M.; van Lohuizen, M.; Vidal, M.; et al. Polycomb function during oogenesis is required for mouse embryonic development. Genes Dev. 2012, 26, 920–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akasaka, T.; Kanno, M.; Balling, R.; Mieza, M.A.; Taniguchi, M.; Koseki, H. A role for mel-18, a Polycomb group-related vertebrate gene, during theanteroposterior specification of the axial skeleton. Development 1996, 122, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- van der Lugt, N.M.; Domen, J.; Linders, K.; van Roon, M.; Robanus-Maandag, E.; te Riele, H.; van der Valk, M.; Deschamps, J.; Sofroniew, M.; van Lohuizen, M.; et al. Posterior transformation, neurological abnormalities, and severe hematopoietic defects in mice with a targeted deletion of the bmi-1 proto-oncogene. Genes Dev. 1994, 8, 757–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.; Pintacuda, G.; Masui, O.; Koseki, Y.; Gdula, M.; Cerase, A.; Brown, D.; Mould, A.; Innocent, C.; Nakayama, M.; et al. PCGF3/5-PRC1 initiates Polycomb recruitment in X chromosome inactivation. Science 2017, 356, 1081–1084. [Google Scholar] [CrossRef]
- Endoh, M.; Endo, T.A.; Shinga, J.; Hayashi, K.; Farcas, A.; Ma, K.W.; Ito, S.; Sharif, J.; Endoh, T.; Onaga, N.; et al. PCGF6-PRC1 suppresses premature differentiation of mouse embryonic stem cells by regulating germ cell-related genes. Elife 2017, 6, e21064. [Google Scholar] [CrossRef]
- Qin, J.; Whyte, W.A.; Anderssen, E.; Apostolou, E.; Chen, H.H.; Akbarian, S.; Bronson, R.T.; Hochedlinger, K.; Ramaswamy, S.; Young, R.A.; et al. The polycomb group protein L3mbtl2 assembles an atypical PRC1-family complex that is essential in pluripotent stem cells and early development. Cell Stem Cell 2012, 11, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Carroll, D.; Erhardt, S.; Pagani, M.; Barton, S.C.; Surani, M.A.; Jenuwein, T. The polycomb-group gene Ezh2 is required for early mouse development. Mol. Cell. Biol. 2001, 21, 4330–4336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, C.; Schumacher, A.; Holdener, B.; Magnuson, T. The eed mutation disrupts anterior mesoderm production in mice. Development 1995, 121, 273–285. [Google Scholar] [CrossRef]
- Pasini, D.; Bracken, A.P.; Jensen, M.R.; Lazzerini Denchi, E.; Helin, K. Suz12 is essential for mouse development and for EZH2 histone methyltransferase activity. EMBO J. 2004, 23, 4061–4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrup, O.; Reiner, A.H.; Aleström, P.; Collas, P. The specific alteration of histone methylation profiles by DZNep during early zebrafish development. Biochim. Biophys. Acta 2014, 1839, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Schultz-Rogers, L.E.; Thayer, M.L.; Kambakam, S.; Wierson, W.A.; Helmer, J.A.; Wishman, M.D.; Wall, K.A.; Greig, J.L.; Forsman, J.L.; Puchhalapalli, K.; et al. Rbbp4 loss disrupts neural progenitor cell cycle regulation independent of Rb and leads to Tp53 acetylation and apoptosis. Dev. Dyn. 2022, 251, 1267–1290. [Google Scholar] [CrossRef] [PubMed]
- Ezhkova, E.; Lien, W.H.; Stokes, N.; Pasolli, H.A.; Silva, J.M.; Fuchs, E. EZH1 and EZH2 cogovern histone H3K27 trimethylation and are essential for hair follicle homeostasis and wound repair. Genes Dev. 2011, 25, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, W.K.; Kang, K.; Yu, J.H.; Yoo, K.H.; Factor, V.M.; Kaji, K.; Matter, M.; Thorgeirsson, S.; Hennighausen, L. The methyltransferases enhancer of zeste homolog (EZH) 1 and EZH2 control hepatocyte homeostasis and regeneration. FASEB J. 2015, 29, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Li, G.; Sarma, K.; Blais, A.; Zavadil, J.; Woodcock, C.L.; Dynlacht, B.D.; Reinberg, D. Ezh1 and Ezh2 maintain repressive chromatin through different mechanisms. Mol. Cell 2008, 32, 503–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavarone, E.; Barbieri, C.M.; Pasini, D. Dissecting the role of H3K27 acetylation and methylation in PRC2 mediated control of cellular identity. Nat. Commun. 2019, 10, 1679. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Liu, Y.; Hsu, Y.J.; Fujiwara, Y.; Kim, J.; Mao, X.; Yuan, G.C.; Orkin, S.H. EZH1 mediates methylation on histone H3 lysine 27 and complements EZH2 in maintaining stem cell identity and executing pluripotency. Mol. Cell 2008, 32, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Holder, M.; Grau, D.; Saldaña-Meyer, R.; Yu, J.R.; Ganai, R.A.; Zhang, J.; Wang, M.; LeRoy, G.; Dobenecker, M.W.; et al. Distinct Stimulatory Mechanisms Regulate the Catalytic Activity of Polycomb Repressive Complex 2. Mol. Cell 2018, 70, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Zhang, Y. SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED-EZH2 complex. Mol. Cell 2004, 15, 57–67. [Google Scholar] [CrossRef]
- Ferrari, K.J.; Scelfo, A.; Jammula, S.; Cuomo, A.; Barozzi, I.; Stützer, A.; Fischle, W.; Bonaldi, T.; Pasini, D. Polycomb-dependent H3K27me1 and H3K27me2 regulate active transcription and enhancer fidelity. Mol. Cell 2014, 53, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, M.; Sugimoto, K.; Fukushima, T.; Shinkai, Y. Set domain-containing protein, G9a, is a novel lysine-preferring mammalian histone methyltransferase with hyperactivity and specific selectivity to lysines 9 and 27 of histone H3. J. Biol. Chem. 2001, 276, 25309–25317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachibana, M.; Ueda, J.; Fukuda, M.; Takeda, N.; Ohta, T.; Iwanari, H.; Sakihama, T.; Kodama, T.; Hamakubo, T.; Shinkai, Y. Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9. Genes Dev. 2005, 19, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Chen, X.; Xiong, J.; Li, Y.; Li, H.; Ding, X.; Liu, S.; Chen, S.; Gao, S.; Zhu, B. Histone methyltransferase G9a contributes to H3K27 methylation in vivo. Cell Res. 2011, 21, 365–367. [Google Scholar] [CrossRef] [Green Version]
- Rougeot, J.; Chrispijn, N.D.; Aben, M.; Elurbe, D.M.; Andralojc, K.M.; Murphy, P.J.; Jansen, P.W.T.C.; Vermeulen, M.; Cairns, B.R.; Kamminga, L.M. Maintenance of spatial gene expression by Polycomb-mediated repression after formation of a vertebrate body plan. Development 2019, 146, dev178590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, G.J.; Wike, C.L.; Nie, X.; Guo, Y.; Tan, M.; Murphy, P.J.; Cairns, B.R. Establishment of developmental gene silencing by ordered polycomb complex recruitment in early zebrafish embryos. Elife 2022, 11, e67738. [Google Scholar] [CrossRef]
- San, B.; Aben, M.; Elurbe, D.M.; Voeltzke, K.; Den Broeder, M.J.; Rougeot, J.; Legler, J.; Kamminga, L.M. Genetic and Epigenetic Regulation of Zebrafish Intestinal Development. Epigenomes 2018, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Lucore, E.C.; Connaughton, V.P. Observational learning and irreversible starvation in first-feeding zebrafish larvae: Is it okay to copy from your friends? Zoology 2021, 145, 125896. [Google Scholar] [CrossRef] [PubMed]
- Chrispijn, N.D.; Elurbe, D.M.; Mickoleit, M.; Aben, M.; de Bakker, D.E.M.; Andralojc, K.M.; Huisken, J.; Bakkers, J.; Kamminga, L.M. Loss of the Polycomb group protein Rnf2 results in derepression of tbx-transcription factors and defects in embryonic and cardiac development. Sci. Rep. 2019, 9, 4327. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.T.; Kathrein, K.L.; Barton, A.; Gitlin, Z.; Huang, Y.H.; Ward, T.P.; Hofmann, O.; Dibiase, A.; Song, A.; Tyekucheva, S.; et al. A network of epigenetic regulators guides developmental haematopoiesis in vivo. Nat. Cell Biol. 2013, 15, 1516–1525. [Google Scholar] [CrossRef]
- Feng, G.; Sun, Y. The Polycomb group gene rnf2 is essential for central and enteric neural system development in zebrafish. Front. Neurosci. 2022, 16, 960149. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.L.; Morey, L. Emerging Roles for Polycomb-Group Proteins in Stem Cells and Cancer. Trends Biochem. Sci. 2019, 44, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Fazio, M.; Ablain, J.; Chuan, Y.; Langenau, D.M.; Zon, L.I. Zebrafish patient avatars in cancer biology and precision cancer therapy. Nat. Rev. Cancer 2020, 20, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Raby, L.; Völkel, P.; Le Bourhis, X.; Angrand, P.O. Genetic Engineering of Zebrafish in Cancer Research. Cancers 2020, 12, 2168. [Google Scholar] [CrossRef]
- Weiss, J.M.; Lumaquin-Yin, D.; Montal, E.; Suresh, S.; Leonhardt, C.S.; White, R.M. Shifting the focus of zebrafish toward a model of the tumor microenvironment. Elife 2022, 11, e69703. [Google Scholar] [CrossRef]
- Gjini, E.; Jing, C.B.; Nguyen, A.T.; Reyon, D.; Gans, E.; Kesarsing, M.; Peterson, J.; Pozdnyakova, O.; Rodig, S.J.; Mansour, M.R.; et al. Disruption of asxl1 results in myeloproliferative neoplasms in zebrafish. Dis. Model Mech. 2019, 12, dmm035790. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Xu, S.; Zhang, Y.; Xu, J.; Huang, Z.; Liu, W.; Wang, S.; Yen, K.; Zhang, W. Asxl1 C-terminal mutation perturbs neutrophil differentiation in zebrafish. Leukemia 2021, 35, 2299–2310. [Google Scholar] [CrossRef]
- Fisher, C.L.; Randazzo, F.; Humphries, R.K.; Brock, H.W. Characterization of Asxl1, a murine homolog of Additional sex combs, and analysis of the Asx-like gene family. Gene 2006, 369, 109–118. [Google Scholar] [CrossRef]
- Kolovos, P.; Nishimura, K.; Sankar, A.; Sidoli, S.; Cloos, P.A.; Helin, K.; Christensen, J. PR-DUB maintains the expression of critical genes through FOXK1/2- and ASXL1/2/3-dependent recruitment to chromatin and H2AK119ub1 deubiquitination. Genome Res. 2020, 30, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Reddington, C.J.; Fellner, M.; Burgess, A.E.; Mace, P.D. Molecular Regulation of the Polycomb Repressive-Deubiquitinase. Int. J. Mol. Sci. 2020, 21, 7837. [Google Scholar] [CrossRef]
- Abdel-Wahab, O.; Pardanani, A.; Patel, J.; Wadleigh, M.; Lasho, T.; Heguy, A.; Beran, M.; Gilliland, D.G.; Levine, R.L.; Tefferi, A. Concomitant analysis of EZH2 and ASXL1 mutations in myelofibrosis, chronic myelomonocytic leukemia and blast-phase myeloproliferative neoplasms. Leukemia 2011, 25, 1200–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boultwood, J.; Perry, J.; Pellagatti, A.; Fernandez-Mercado, M.; Fernandez-Santamaria, C.; Calasanz, M.J.; Larrayoz, M.J.; Garcia-Delgado, M.; Giagounidis, A.; Malcovati, L.; et al. Frequent mutation of the polycomb-associated gene ASXL1 in the myelodysplastic syndromes and in acute myeloid leukemia. Leukemia 2010, 24, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Gelsi-Boyer, V.; Trouplin, V.; Roquain, J.; Adélaïde, J.; Carbuccia, N.; Esterni, B.; Finetti, P.; Murati, A.; Arnoulet, C.; Zerazhi, H.; et al. ASXL1 mutation is associated with poor prognosis and acute transformation in chronic myelomonocytic leukaemia. Br. J. Haematol. 2010, 151, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelsi-Boyer, V.; Brecqueville, M.; Devillier, R.; Murati, A.; Mozziconacci, M.J.; Birnbaum, D. Mutations in ASXL1 are associated with poor prognosis across the spectrum of malignant myeloid diseases. J. Hematol. Oncol. 2012, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, D.; Kitaura, J.; Togami, K.; Nishimura, K.; Enomoto, Y.; Uchida, T.; Kagiyama, Y.; Kawabata, K.C.; Nakahara, F.; Izawa, K.; et al. Myelodysplastic syndromes are induced by histone methylation–altering ASXL1 mutations. J. Clin. Investig. 2013, 123, 4627–4640. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; You, X.; Droin, N.; Banaszak, L.G.; Churpek, J.; Padron, E.; Geissler, K.; Solary, E.; Patnaik, M.M.; Zhang, J. Role of ASXL1 in hematopoiesis and myeloid diseases. Exp. Hematol. 2022, 115, 14–19. [Google Scholar] [CrossRef]
- Geng, Y.; Zhang, T.; Alonzo, I.G.; Godar, S.C.; Yates, C.; Pluimer, B.R.; Harrison, D.L.; Nath, A.K.; Yeh, J.J.; Drummond, I.A.; et al. Top2a promotes the development of social behavior via PRC2 and H3K27me3. Sci. Adv. 2022, 8, eabm7069. [Google Scholar] [CrossRef]


| Complex | Complex Type | Subunit | Zebrafish Ortholog |
|---|---|---|---|
| PRC2 | Core | EZH2 EZH1 EED SUZ12 RBBP4 RBBP7 | Ezh2 Ezh1 Eed Suz12a, Suz12b Rbbp4 - 1 |
| PRC2 | PRC2.1 | EPOP PAL1 (LCOR) PAL2 (LCORL) PHF1 MTF2 PH19 | Skida1l 2 Lcor Lcorl Phf1 Mtf2 Phf19 |
| PRC2 | PRC2.2 | AEBP2 JARID2 | Aebp2 Jarid2a, Jarid2b |
| PRC1 | Core | PCGF1 PCGF2 PCGF3 BMI1 (PCGF4) PCGF5 PCGF6 RNF2 RING1 | Pcgf1 - - Bmi1a, Bmi1b Pcgf5a, Pcgf5b Pcgf6 Rnf2 - |
| PRC1 | cPRC1 | CBX2 CBX4 CBX6 CBX7 CBX8 PHC1 PHC2 PHC3 | Cbx2 Cbx4 Cbx6a, Cbx6b Cbx7a, Cbx7b Cbx8a, Cbx8b Phc1 Phc2a, Phc2b Phc3 |
| PRC1 | ncPRC1 | RYBP YAF2 BCOR BCORL1 KDM2 USP7 SKP1 AUTS2 FBRS FBRSL1 E2F6 CBX3 HDAC1 HDAC2 L3MBTL2 WDR5 MAX MGA | Rybpa, Rybpb Yaf2 Bcor Bcol1 Kdm2ba, Kdm2bb Usp7 Skp1 Auts2a, Auts2b Fbrs Fbrsl1 E2f6 Cbx3a, Cbx3b Hdac1 - L3mbtl2 Wdr5 Max Mgaa, Mgab |
| Subunit | Zebrafish | Medaka | Japanese Pufferfish |
|---|---|---|---|
| RING1 RNF2 PCGF1 PCGF2 PCGF3 BMI1 (PCGF4) PCGF5 PCGF6 | − + + − − ++ ++ + | + + − + + + + + | − + + + − + + + |
| Subunit | Zebrafish | Medaka | Japanese Pufferfish |
|---|---|---|---|
| CBX2 CBX4 CBX6 CBX7 CBX8 | + + ++ ++ ++ | ++ ++ + ++ ++ | ++ ++ + ++ ++ |
| Gene | Technique | Allele | References |
|---|---|---|---|
| ezh1 | MO TALEN CRISPR/Cas9 | - ul3 b1394 | [68] [69] [70] |
| ezh2 | MO ENU TALEN ENU CRISPR/Cas9 | - hu5670 ul2 sa1199 2 b1392 | [71] [72] [73] [70,71,74] [70] |
| eed | TALEN | ul4 | [75] |
| suz12a/b | CRISPR/Cas9 | [76] | |
| rbbp4 | MO CRISPR/Cas9 | - is60 | [77] [78] |
| rnf2 | MO ZFN CRISPR/Cas9 | - ibl31 f5, f8 | [79] [80] [81] |
| pcgf1 | MO TALEN | - ul1 | [82] [83] |
| bmi1a/b | MO | - | [84,85] |
| phc1 | MO | - | [86] |
| phc2a | MO | - | [87] |
| yaf2 | MO | - | [88] |
| bcor | MO | - | [89,90] |
| kdm2bb | MO | - | [91] |
| usp7 | MO | - | [92] |
| skp1 | IM 1 | hi3970Tg | [93] |
| wdr5 | CRISPR/Cas9 | zju131 | [94] |
| auts2a | TALEN | ncb104 | [95] |
| mgaa | MO TALEN | - ihb801 | [96] [97] |
| max | MO | - | [98] |
| hdac1 | MO IM | - hi1618Tg | [99] [99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanot, M.; Raby, L.; Völkel, P.; Le Bourhis, X.; Angrand, P.-O. The Contribution of the Zebrafish Model to the Understanding of Polycomb Repression in Vertebrates. Int. J. Mol. Sci. 2023, 24, 2322. https://doi.org/10.3390/ijms24032322
Hanot M, Raby L, Völkel P, Le Bourhis X, Angrand P-O. The Contribution of the Zebrafish Model to the Understanding of Polycomb Repression in Vertebrates. International Journal of Molecular Sciences. 2023; 24(3):2322. https://doi.org/10.3390/ijms24032322
Chicago/Turabian StyleHanot, Mariette, Ludivine Raby, Pamela Völkel, Xuefen Le Bourhis, and Pierre-Olivier Angrand. 2023. "The Contribution of the Zebrafish Model to the Understanding of Polycomb Repression in Vertebrates" International Journal of Molecular Sciences 24, no. 3: 2322. https://doi.org/10.3390/ijms24032322