
Insecticidal and Detoxification Enzyme Inhibition Activities of Essential Oils for the Control of Pulse Beetle, Callosobruchus maculatus (F.) and Callosobruchus chinensis (L.) (Coleoptera: Bruchidae)


Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of Essential Oils
2.2. Fumigant Toxicity of Essential Oils against C. maculatus and C. chinensis
2.2.1. C. maculatus
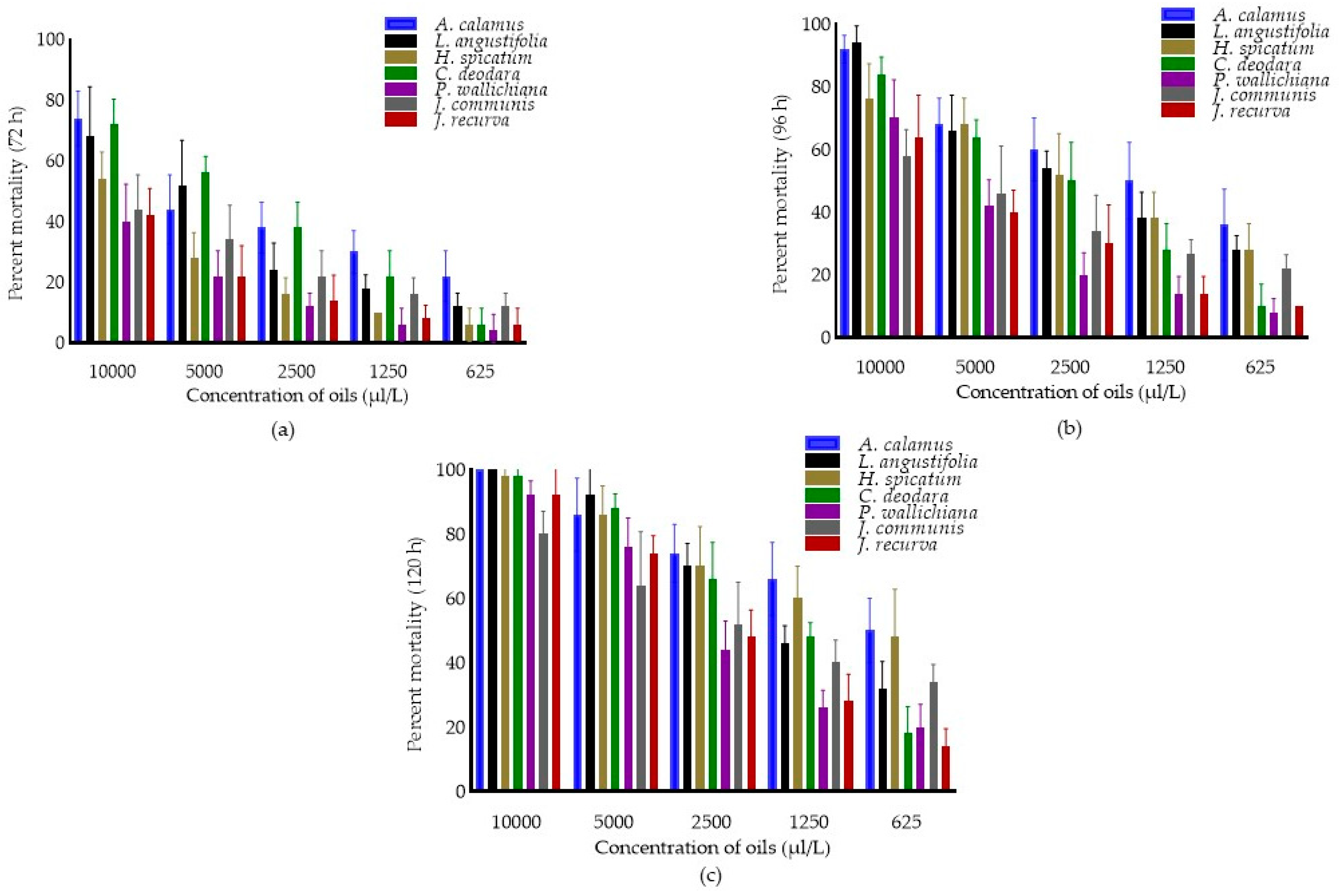
Toxicity of Different Essential Oils against C. maculatus
Percent Mortality of Essential Oils against C. maculatus
2.2.2. C. chinensis
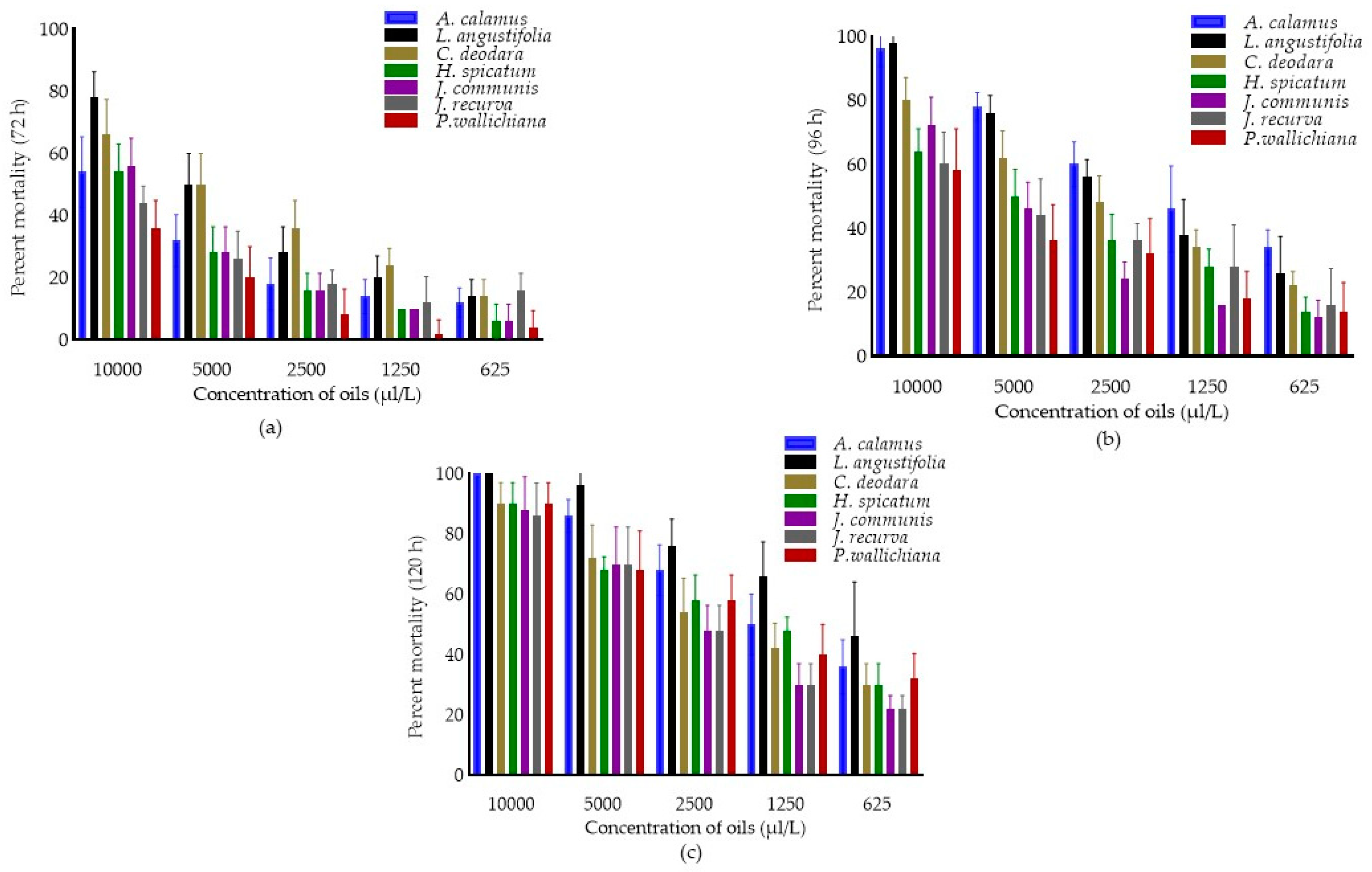
Toxicity of Different Essential Oils against C. chinensis
Percent Mortality of Different EOs against C. chinensis
2.3. Repellent Activity of Essential Oils against Pulse Beetle
2.4. Ovipositional Inhibition (OI) Effect of Essential Oils against Pulse Beetle
2.4.1. C. maculatus
2.4.2. C. chinensis
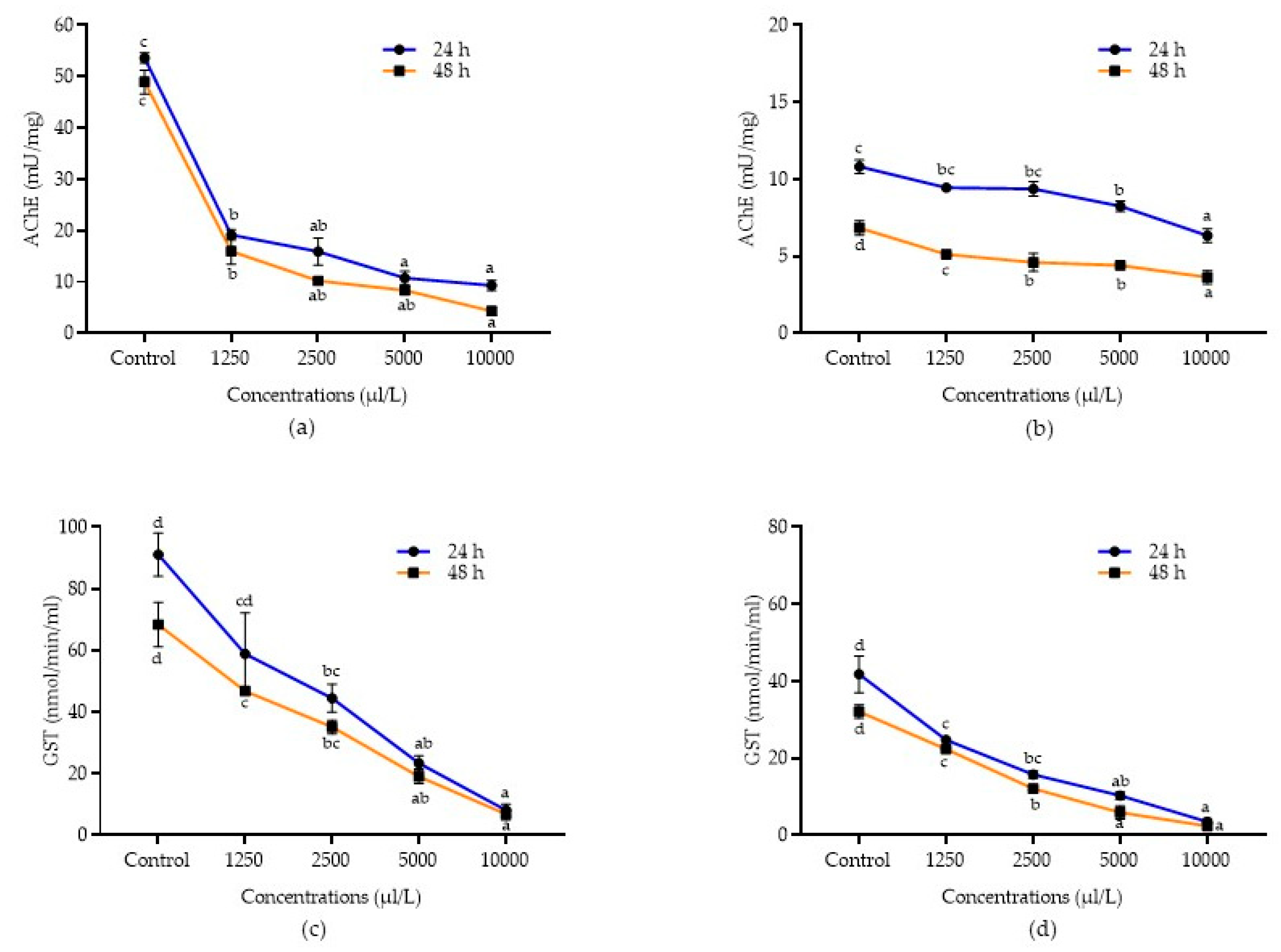
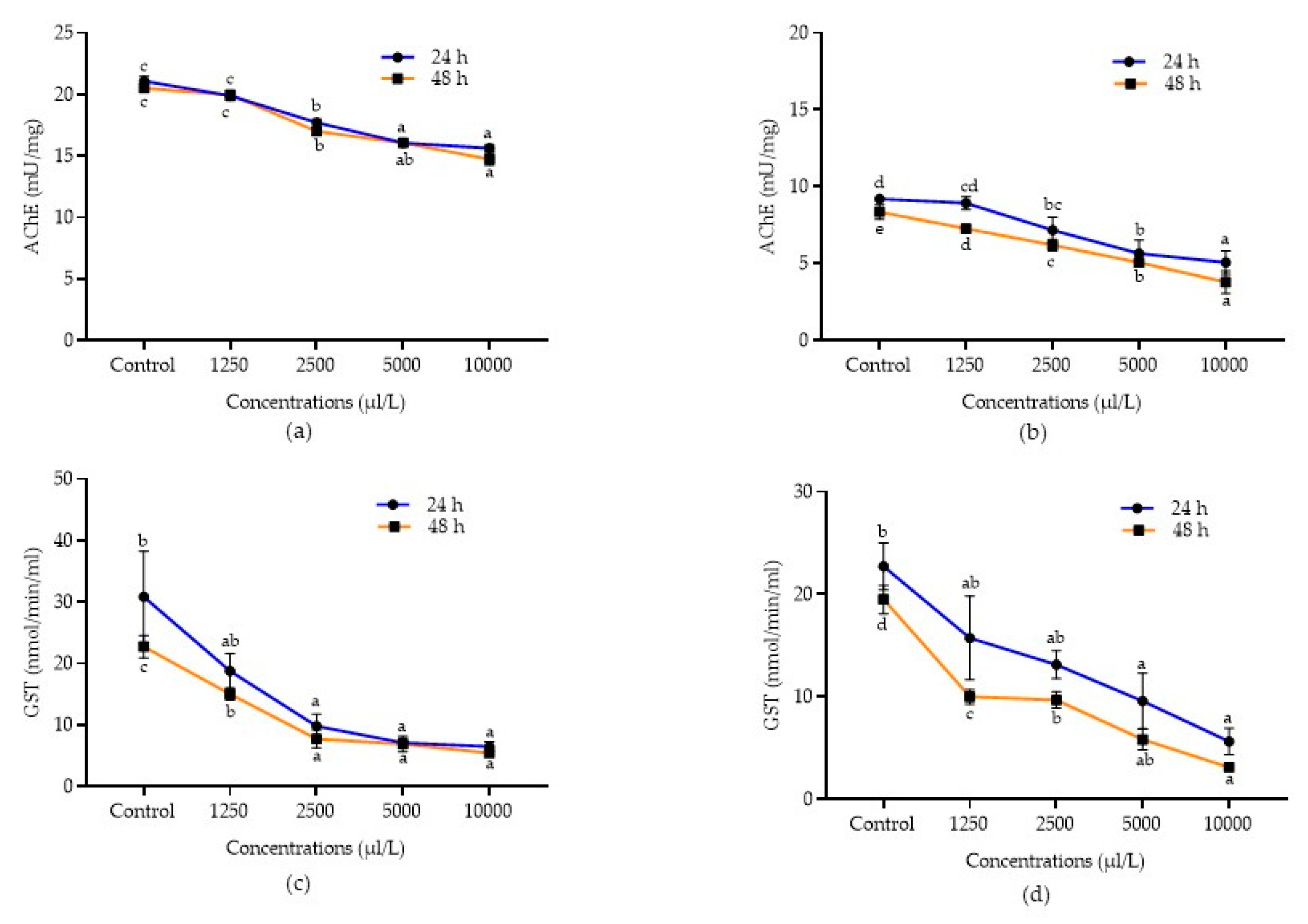
2.5. Detoxification Enzyme Activities of A. Calamus and L. Angustifolia against Pulse Beetle
3. Discussion
4. Materials and Methods
4.1. Essential Oils (EOs)
4.2. Test Insect
4.3. Gas Chromatography (GC) Analysis
4.4. GC-MS Analysis and Identification
4.5. Fumigant Toxicity of Essential Oils and Their Combinations against the Pulse Beetle
4.6. Repellent Activity of Essential Oils against the Pulse Beetle
4.7. Ovipositional Deterrent Activity of Essential Oils against the Pulse Beetle
4.8. Detoxification Enzyme Inhibition of A. Calamus and L. Angustifolia EO against the Pulse Beetle
4.8.1. Sample Preparation
4.8.2. Protein Estimation
4.8.3. Acetylcholinesterase Assay
4.8.4. Glutathione-S-Transferase Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bessada, S.M.; Barreira, J.C.; Oliveira, M.B.P. Pulses and food security: Dietary protein, digestibility, bioactive and functional properties. Trends Food Sci. Technol. 2019, 93, 53–68. [Google Scholar] [CrossRef]
- Murugesan, R.; Vasuki, K.; Kaleeswaran, B.; Santhanam, P.; Ravikumar, S.; Alwahibi, M.S.; Soliman, D.A.; Mohsen Ahmed Almunqedhi, B.; Alkahtani, J. Insecticidal and repellent activities of Solanum torvum (Sw.) leaf extract against stored grain Pest, Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. King Saud Univ.-Sci. 2021, 33, 101390. [Google Scholar] [CrossRef]
- Seni, A.; Mishra, K.M. Pulse beetle, Callosobruchus spp. (Coleoptera: Chrysomelidae); a major threat in legume grain storage and their management. Acta Phytopathol. Entomol. Hung. 2022, 57, 49–65. [Google Scholar] [CrossRef]
- Mogbo, T.C.; Okeke, T.E.; Akunne, C.E. Studies on the resistance of cowpea seeds (Vigna unguiculata) to weevil (Callosobruchus maculatus) infestations. Am. J. Zool. Res. 2014, 2, 37–40. [Google Scholar] [CrossRef]
- Tuda, M.; Chou, L.Y.; Niyomdham, C.; Buranapanichpan, S.; Tateishi, Y. Ecological factors associated with pest status in Callosobruchus (Coleoptera: Bruchidae): High host specificity of non-pests to Cajaninae (Fabaceae). J. Stored Prod. Res. 2005, 41, 31–45. [Google Scholar] [CrossRef]
- Ajayi, O. Bioactivity of the leaf extracts of Morinda lucida (Benth.) against cowpea Bruchid, Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae). Exp. Agric. Hortic. 2012, 1, 1–7. [Google Scholar]
- Beck, C.W.; Blumer, L.S. A Handbook on Bean Beetles, Callosobruchus maculatus; National Science Foundation: Alexandria, VA, USA, 2014; Available online: http://www.beanbeetles.org/handbook.pdf (accessed on 3 October 2022).
- França, A.F.J.; Araújo, J.N.; Santos, Y.Q.; Carelli, G.S.C.; Silva, D.A.; Amorim, T.M.L.; Migliolo, L.; Santos, E.A.; Oliveira, A.S.; Uchoa, A.F. Vicilin from Anadenanthera colubrina Seeds: An alternative tool to combat Callosobruchus maculatus. Saudi J. Biol. Sci. 2021, 28, 5229–5237. [Google Scholar] [CrossRef]
- Jayaram, C.S.; Chauhan, N.; Dolma, S.K.; Reddy, S.G.E. Chemical composition, and insecticidal activities of essential oils against the pulse beetle. Molecules 2022, 27, 568. [Google Scholar] [CrossRef] [PubMed]
- Garry, V.F.; Griffith, J.; Danzl, T.J.; Nelson, R.L.; Whorton, E.B.; Krueger, L.A.; Cervenka, J. Human genotoxicity: Pesticide applicators and phosphine. Science 1989, 246, 251–255. [Google Scholar] [CrossRef]
- Okonkwo, E.U.; Okoye, W.I. The efficacy of four seed powders and the essential oils as protectants of cowpea and maize grains against infestation by Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae) and Sitophilus zeamais (Motschulsky) (Coleoptera: Curculionidae) in Nigeria. Int. J. Pest Manag. 1996, 42, 143–146. [Google Scholar] [CrossRef]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E.E. Repellent activity of essential oils from seven aromatic plants grown in Colombia against Sitophilus zeamais Motschulsky (Coleoptera). J. Stored Prod. Res. 2009, 45, 212–214. [Google Scholar] [CrossRef]
- Vendan, S.E.; Manivannan, S.; Sunny, A.M.; Murugesan, R. Phytochemical residue profiles in rice grains fumigated with essential oils for the control of rice weevil. PLoS ONE 2017, 12, e0186020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.M.; de Freitas Rocha, J.; de Almeida, T.S.; Mota, E.F. Essential oils of caatinga plants with deletary action for Aedes aegypti: A Review. S. Afr. J. Bot. 2021, 143, 69–78. [Google Scholar] [CrossRef]
- Rajendran, S.; Sriranjini, V. Plant products as fumigants for stored-product insect control. J. Stored Prod. Res. 2008, 44, 126–135. [Google Scholar] [CrossRef]
- Rajashekar, Y.; Bakthavatsalam, N.; Shivanandappa, T. Botanicals as Grain Protectants. Psyche 2012, e646740. [Google Scholar] [CrossRef]
- Singh, K.D.; Mobolade, A.J.; Bharali, R.; Sahoo, D.; Rajashekar, Y. Main plant volatiles as stored grain pest management approach: A review. J. Agric. Food Res. 2021, 4, 100127. [Google Scholar] [CrossRef]
- Dubey, V.S.; Bhalla, R.; Luthra, R. An overview of the non-mevalonate pathway for terpenoid biosynthesis in plants. J. Biosci. 2003, 28, 637–646. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Ann. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Chauhan, N.; Kashyap, U.; Dolma, S.K.; Reddy, S.G.E. Chemical composition, insecticidal, persistence and detoxification enzyme inhibition activities of essential oil of Artemisia maritima against the pulse beetle. Molecules 2022, 27, 1547. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured Publication: Carol Stream, IL, USA, 2007. [Google Scholar]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.O.L.C.; Obame, L.C.; Ilboudo, A.J.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Chemical composition and antimicrobial activity of Cymbopogon citratus and Cymbopogon giganteus essential oils alone and in combination. Phytomedicine 2011, 18, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Guccione, C.; Isacchi, B.; Righeschi, C.; Firenzuoli, F.; Bergonzi, M.C. Essential oils loaded in nanosystems: A developing strategy for a successful therapeutic approach. Evid.-Based Complement. Altern. Med. 2014, 2014, 651593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perricone, M.; Arace, E.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Bioactivity of essential oils: A review on their interaction with food components. Front. Microbiol. 2015, 6, 76. [Google Scholar] [CrossRef] [Green Version]
- Hanif, M.A.; Nisar, S.; Khan, G.S.; Mushtaq, Z.; Zubair, M. Essential oils. In Essential Oil Research; Malik, S., Ed.; Springer: Cham, Switzerland, 2019; pp. 3–17. [Google Scholar] [CrossRef]
- Kim, S.I.; Roh, J.Y.; Kim, D.H.; Lee, H.S.; Ahn, Y.J. Insecticidal activities of aromatic plant extracts and essential oils against Sitophilus oryzae and Callosobruchus chinensis. J. Stored Prod. Res. 2003, 39, 293–303. [Google Scholar] [CrossRef]
- Kalagatur, N.K.; Kamasani, J.R.; Siddaiah, C.; Gupta, V.K.; Krishna, K.; Mudili, V. Combinational inhibitory action of Hedychium spicatum L. essential oil and γ-radiation on growth rate and mycotoxins content of Fusarium graminearum in maize: Response surface methodology. Front. Microbiol. 2018, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Rawat, A.; Thapa, P.; Prakash, O.; Kumar, R.; Pant, A.K.; Srivastava, R.M.; Rawat, D.S. Chemical composition, herbicidal, antifeedant and cytotoxic activity of Hedychium spicatum Sm.: A Zingiberaceae herb. Trends Phytochem. Res. 2019, 3, 123–136. [Google Scholar]
- Arya, S.; Kumar, R.; Prakash, O.; Kumar, S.; Mahawer, S.K.; Chamoli, S.; de Oliveira, M.S. Chemical composition and biological activities of Hedychium coccineum Buch.-Ham. ex Sm. essential oils from Kumaun hills of Uttarakhand. Molecules 2022, 27, 4833. [Google Scholar] [CrossRef]
- Miao, J.K.; Shi, R.H.; Li, C.; Li, X.W.; Chen, Q.X. Sweet Flag (Acorus calamus) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2016; pp. 775–782. [Google Scholar] [CrossRef]
- Brito, V.D.; Achimón, F.; Pizzolitto, R.P.; Ramírez Sánchez, A.; Gómez Torres, E.A.; Zygadlo, J.A.; Zunino, M.P. An alternative to reduce the use of the synthetic insecticide against the maize weevil Sitophilus zeamais through the synergistic action of Pimenta racemosa and Citrus sinensis essential oils with chlorpyrifos. J. Pest Sci. 2021, 94, 409–421. [Google Scholar] [CrossRef]
- Singh, P.; Andola, H.; Rawat, M.S.M.; Jangwan, J.S. GC-MS analysis of essential oil from Lavandula angustifolia cultivated in Garhwal Himalaya. J. Nat. Prod. 2015, 5, 268–272. [Google Scholar] [CrossRef]
- Beale, D.J.; Morrison, P.D.; Karpe, A.V.; Dunn, M.S. Chemometric analysis of lavender essential oils using targeted and untargeted GC-MS acquired data for the rapid identification and characterization of oil quality. Molecules 2017, 22, 1339. [Google Scholar] [CrossRef]
- Dong, G.; Bai, X.; Aimila, A.; Aisa, H.A.; Maiwulanjiang, M. Study on lavender essential oil chemical compositions by GC-MS and improved pGC. Molecules 2020, 25, 3166. [Google Scholar] [CrossRef] [PubMed]
- Dambolena, J.S.; Gallucci, M.N.; Luna, A.; Gonzalez, S.B.; Guerra, P.E.; Zunino, M.P. Composition, antifungal and antifumonisin activity of Pinus wallichiana, Pinus monticola and Pinus strobus essential oils from Patagonia Argentina. J. Essent. Oil-Bear. Plants 2016, 19, 1769–1775. [Google Scholar] [CrossRef]
- Rashed, K. Phytochemical and biological activities of Pinus wallichiana: A short review. Int. J. Pharm. Sci. Drug Anal. 2022, 2, 38–40. [Google Scholar]
- Kumar, S.; Mitra, B.; Kashyap, S.; Kumar, S. Physicochemical properties, GC-MS analysis and impact of different material size on yield of Himalayan C. deodara essential oil. Pharmacogn. Res. 2022, 14, 181–187. [Google Scholar] [CrossRef]
- Perry, N.B.; Anderson, R.E.; Brennan, N.J.; Douglas, M.H.; Heaney, A.J.; McGimpsey, J.A.; Smallfield, B.M. Essential oils from dalmatian sage (Salvia officinalis L.): Variations among individuals, plant parts, seasons, and sites. J. Agric. Food Chem. 1999, 47, 2048–2054. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, B.H.; Choi, W.S.; Park, B.S.; Kim, J.G.; Campbell, B.C. Fumigant toxicity of volatile natural products from Korean spices and medicinal plants towards the rice weevil, Sitophilus oryzae (L). Pest Manag. Sci. 2001, 57, 548–553. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Prajapati, V.; Aggarwal, K.K.; Kumar, S. Toxicity, feeding deterrence, and effect of the activity of 1, 8-cineole from Artemisia annua on progeny production of Tribolium castanaeum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2001, 94, 979–983. [Google Scholar] [CrossRef]
- Hafez, M.; Dimetry, N.Z.; Abbass, M.H. Insecticidal and biological efficacy of two vegetable oils against Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae) under laboratory conditions. Arch. Phytopathol. Pflanzenschutz. 2014, 47, 1942–1955. [Google Scholar] [CrossRef]
- Baskaran, J.; Dhanasekaran, S.; Krishnappa, K.; Elumalai, K. Toxicity of essential oils used for the protection of stored cowpea against Callosobruchus maculatus F. (Leoptera: Bruchidae). Int. J. Curr. Res. 2010, 9, 039–045. [Google Scholar]
- Su, H.C. Laboratory evaluation of toxicity of calamus oil against four species of stored-product insects. J. Entomol. Sci. 1991, 26, 76–80. [Google Scholar] [CrossRef]
- Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Assessment of essential oil of Acorus calamus L. and its major constituent β-Asarone in post-harvest management of Callosobruchus chinensis L. Acta Phytopathol. Entomol. Hung. 2016, 19, 542–552. [Google Scholar] [CrossRef]
- Hasan, M.S.; Ullah, E.; Ahmad, F.; Wakil, W. Insecticidal activity of different doses of Acorus calamus oil against Trogoderma granarium (Everts). Pak. J. Agric. Sci. 2006, 43, 55–58. [Google Scholar]
- Koundal, R.; Dolma, S.K.; Chand, G.; Agnihotri, V.K.; Reddy, S.G.E. Chemical composition, and insecticidal properties of essential oils against diamondback moth (Plutella xylostella L.). Toxin Rev. 2020, 39, 371–381. [Google Scholar] [CrossRef]
- Sakhanokho, H.F.; Sampson, B.J.; Tabanca, N.; Wedge, D.E.; Demirci, B.; Baser, K.H.C.; Spiers, J.M. Chemical composition, antifungal and insecticidal activities of Hedychium essential oils. Molecules 2013, 18, 4308–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruda, M.; Viana, H.; Rainha, N.; Neng, N.R.; Rosa, J.S.; Nogueira, J.M.; Barreto, M.D.C. Anti-acetylcholinesterase and antioxidant activity of essential oils from Hedychium gardnerianum Sheppard ex Ker-Gawl. Molecules 2012, 17, 3082–3092. [Google Scholar] [CrossRef] [Green Version]
- Ncibi, S.; Barbouche, N.; Haouel-hamdi, S.; Ammar, M. Insecticidal activity of several Tunisian essential oils against two major pests of stored grain Rhyzopertha dominica (Fabricius, 1792) and Tribolium castaneum (Herbest 1797). J. New Sci. Agric. Biotech. 2019, 66, 4182–4194. [Google Scholar]
- Sayada, N.; Tine, S.; Soltani, N. Evaluation of a botanical insecticide, lavender (Lavandula angustifolia (M.)) essential oil as toxicant, repellent and antifeedant against lesser grain borer (Rhyzopertha dominica (F.)). Appl. Ecol. Environ. Res. 2008, 20, 1301–1324. [Google Scholar] [CrossRef]
- Abdali, E.Y.; Agour, A.; Allali, A.; Bourhia, M.; Moussaoui, E.A.; Eloutassi, N.; Bouia, A. Lavandula dentata L.: Phytochemical analysis, antioxidant, antifungal and insecticidal activities of its essential oil. Plants 2022, 11, 311. [Google Scholar] [CrossRef]
- Mazarin, A.; Nukenine, E.N.; Niu, C.; Vincent, F.V. Synergistic effects of wood ash and essential oil on fecundity, pupal eclosion and adult mortality of Callosobruchus maculatus (Coleoptera: Bruchidae) cowpea seed weevil. Am. J. Exp. Agric. 2016, 11, 1–12. [Google Scholar] [CrossRef]
- Hategekimana, A.; Erler, F. Comparative repellent activity of single, binary, and ternary combinations of plant essential oils and their major components against Sitophilus oryzae L. (Coleoptera: Curculionidae). J. Plant Prot. Res. 2020, 127, 873–881. [Google Scholar] [CrossRef]
- Brito, R.D.C.; Fontes, L.D.S.; da Silva, P.H.S.; Santana, C.D.S. Essential oils from Betula lenta, Cinnamomum cassia, Citrus aurantium var. Amara and Acorus calamus as biopesticides against cowpea weevil. Int. J. Trop. Insect Sci. 2022, 42, 261–268. [Google Scholar] [CrossRef]
- Rotimi, J.; Evbuomwan, C.O. Deterrent effects of citrus peel oils on oviposition and adult emergence of the cowpea weevil, Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Adv. Appl. Sci. Res. 2012, 3, 3545–3550. [Google Scholar]
- Matos, L.F.; da Cruz Lima, E.; de Andrade Dutra, K.; Navarro, D.M.D.A.F.; Alves, J.L.R.; Silva, G.N. Chemical composition, and insecticidal effect of essential oils from Illicium verum and Eugenia caryophyllus on Callosobruchus maculatus in cowpea. Ind. Crops Prod. 2020, 145, 112088. [Google Scholar] [CrossRef]
- Upadhyay, N.; Dwivedy, A.K.; Kumar, M.; Prakash, B.; Dubey, N.K. Essential oils as eco-friendly alternatives to synthetic pesticides for the control of Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Essent. Oil-Bear. Plants 2018, 21, 282–297. [Google Scholar] [CrossRef]
- Hu, Z.D.; Xia, F.E.N.G.; Lin, Q.S.; Chen, H.Y.; Li, Z.Y.; Fei, Y.I.N.; Gao, X.W. Biochemical mechanism of chlorantraniliprole resistance in the diamondback moth, Plutella xylostella Linnaeus. J. Integr. Agric. 2014, 13, 2452–2459. [Google Scholar] [CrossRef]
- Tak, J.H.; Isman, M.B. Metabolism of citral, the major constituent of lemongrass oil, in the cabbage looper, Trichoplusia ni, and effects of enzyme inhibitors on toxicity and metabolism. Pestic. Biochem. Physiol. 2016, 133, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Piao, X.; Zhang, L.; Song, S.; Xu, Y. Ginsenosides from the stems and leaves of Panax ginseng show antifeedant activity against Plutella xylostella. Ind. Crops Prod. 2018, 124, 412–417. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.; Mohamed, M.I.; Shawir, M.S.; Abou-Taleb, H.K. Chemical composition, insecticidal and biochemical effects of essential oils of different plant species from Northern Egypt on the rice weevil, Sitophilus oryzae L. J. Pest Sci. 2016, 89, 219–229. [Google Scholar] [CrossRef]
- Stein, S.E. Mass Spectral Database and Software, version 3.02; US Department of Commerce, National Institute of Standards and Technology: Gaithersburg, MD, USA, 2005.
- Eccles, K.; Powder-George, Y.L.; Mohammed, F.K.; Khan, A. Efficacy of Artocarpus altilis (Parkinson) Fosberg extracts on contact mortality, repellency, oviposition deterrency and fumigant toxicity of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Int. J. Pest Manag. 2019, 65, 72–78. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Name | RI a | RI b | Area (%) | Mode of Identification |
|---|---|---|---|---|---|
| 1 | trans–β–Ocimene | 1032 | 1033 | 0.62 | MS, RI |
| 2 | β–elemene | 1389 | 1390 | 0.51 | MS, RI |
| 3 | Aristolene (calarene) | 1431 | 1435 | 0.51 | MS, RI |
| 4 | trans methyl isoeugenol | 1451 | 1451 | 0.05 | MS, RI |
| 5 | Viridiflorene | 1496 | 1494 | 0.93 | MS, RI |
| 6 | Bicyclo–germacrene | 1500 | 1512 | 1.04 | MS, RI |
| 7 | δ–cadinene | 1522 | 1522 | 1.26 | MS, RI |
| 8 | Kessane | 1529 | 1528 | 0.76 | MS, RI |
| 9 | Cedranone | 1500 | 1504 | 2.93 | MS, RI |
| 10 | α–calacorene | 1544 | 1549 | 0.83 | MS, RI |
| 11 | cis–asarone | 1616 | 1617 | 85.37 | MS, RI |
| 12 | Trans–asarone | 1675 | 1686 | 2.64 | MS, RI |
| 13 | Thujopsanone | 1641 | 1646 | 0.24 | MS, RI |
| 14 | Murrolene | 1779 | 1769 | 0.11 | MS, RI |
| 15 | Methyl palmitate | 1921 | 1927 | 0.04 | MS, RI |
| Unidentified | – | – | 1.52 | ||
| Total | 99.32 | ||||
| Monoterpene hydrocarbons * | 0.67 | ||||
| Oxygenated monoterpene * | 0.0 | ||||
| Phenylpropanoid | 88.01 | ||||
| Sesquiterpene hydrocarbons * | 5.26 | ||||
| Oxygenated sesquiterpene * | 3.93 |
| Essential Oils | LC50 (µL/L) | Confidence Limits (µL/L) | Slope ± SE | Chi-Square | p-Value | Co – Toxicity Coefficient | Interaction Type |
|---|---|---|---|---|---|---|---|
| A. calamus (72 h) | 4128.22 | 2897.88–6763.92 | 1.06 ± 0.20 | 3.74 | 0.29 | – | – |
| 96 h | 1357.86 | 879.60–1859.42 | 1.23 ± 0.21 | 3.56 | 0.31 | – | – |
| 120 h | 701.48 | 401.22–986.06 | 1.49 ± 0.25 | 4.14 | 0.25 | – | – |
| H. spicatum (72 h) | 10,448.32 | 7116.12–20,223.58 | 1.41 ± 0.24 | 1.69 | 0.64 | – | – |
| 96 h | 2177.08 | 1495.50–3067.74 | 1.12 ± 0.20 | 0.21 | 0.97 | – | – |
| 120 h | 806.92 | 483.38–1116.99 | 1.45 ± 0.24 | 2.81 | 0.42 | – | – |
| J. communis (96 h) | 6497.30 | 4012.23–16,957.41 | 0.84 ± 0.20 | 0.41 | 0.94 | – | – |
| 120 h | 1945.44 | 1256.80–2806.47 | 1.03 ± 0.20 | 0.90 | 0.82 | – | – |
| J. recurva (96 h) | 6684.97 | 4877.87–10,740.24 | 1.41 ± 0.22 | 1.42 | 0.70 | – | – |
| 120 h | 2369.76 | 1929.69–2904.90 | 2.04 ± 0.23 | 0.78 | 0.85 | – | – |
| P. wallichiana (96 h) | 5948.84 | 4559.90–8576.71 | 1.65 ± 0.23 | 2.74 | 0.43 | – | – |
| 120 h | 2313.37 | 1862.92–2865.74 | 1.92 ± 0.23 | 4.16 | 0.24 | – | – |
| C. deodara (72 h) | 4116.25 | 3240.30–5491.22 | 1.67 ± 0.22 | 0.85 | 0.84 | – | – |
| 96 h | 2818.88 | 2253.91–3556.77 | 1.80 ± 0.22 | 0.79 | 0.85 | – | – |
| 120 h | 1487.29 | 1202.79–1793.45 | 2.29 ± 0.27 | 1.04 | 0.79 | – | – |
| L. angustifolia (72 h) | 5204.72 | 3923.80–7658.14 | 1.46 ± 0.22 | 2.34 | 0.50 | – | – |
| 96 h | 1876.15 | 1410.29–2419.49 | 1.52 ± 0.22 | 5.01 | 0.17 | – | – |
| 120 h | 1220.93 | 961.43–1488.72 | 2.21 ± 0.27 | 3.35 | 0.34 | – | – |
| AlPO2 (48 h) | 0.075 | 0.068–0.085 | 3.85 ± 0.92 | 0.15 | 0.98 | – | – |
| 72 h | 0.061 | 0.055–0.065 | 5.38 ± 0.97 | 2.44 | 0.49 | – | – |
| 96 h | 0.054 | 0.049–0.058 | 6.57 ± 1.05 | 4.41 | 0.22 | – | – |
| 120 h | 0.050 | 0.044–0.054 | 7.58 ± 1.24 | 2.94 | 0.40 | – | – |
| A. calamus + L. angustifolia (24 h) | 1148.59 | 730.42–2726.61 | 0.93 ± 0.20 | 1.51 | 0.68 | – | – |
| 48 h | 533.72 | 368.26–840.46 | 1.00 ± 0.20 | 0.81 | 0.85 | – | – |
| 72 h | 204.01 | 108.09–303.27 | 0.96 ± 0.20 | 2.09 | 0.55 | 2023.54 | Synergistic |
| 96 h | 108.58 | 59.95–155.01 | 1.45 ± 0.24 | 1.68 | 0.64 | 1250.56 | Synergistic |
| A. calamus + H. spicatum (24 h) | – | – | – | – | – | – | – |
| 48 h | – | – | – | – | – | – | – |
| 72 h | 493.89 | 311.64–827.68 | 0.85 ± 0.19 | 2.31 | 0.51 | 835.86 | Synergistic |
| 96 h | 164.31 | 77.24–250.33 | 0.99 ± 0.20 | 0.93 | 0.82 | 826.40 | Synergistic |
| A. calamus + C. deodara (24 h) | – | – | – | – | – | – | – |
| 48 h | – | – | – | – | – | – | – |
| 72 h | 1626.04 | 864.18–9816.54 | 0.64 ± 0.19 | 0.16 | 0.98 | 253.88 | Synergistic |
| 96 h | 211.08 | 87.04 – 335.46 | 0.86 ± 0.20 | 0.35 | 0.95 | 643.29 | Synergistic |
| A. calamus + P. wallichiana (24 h) | 1967.74 | 1451.44–3038.81 | 1.34 ± 0.22 | 2.78 | 0.43 | – | – |
| 48 h | 1167.83 | 849.94–1684.36 | 1.19 ± 0.20 | 1.31 | 0.73 | – | – |
| 72 h | 615.31 | 409.99–847.10 | 1.18 ± 0.20 | 2.25 | 0.52 | 670.92 | Synergistic |
| 96 h | 169.89 | 64.53–275.61 | 1.23 ± 0.24 | 5.20 | 0.16 | 799.26 | Synergistic |
| A. calamus + J. communis (24 h) | – | – | – | – | – | – | – |
| 48 h | 1798.50 | 940.07–7815.84 | 0.56 ± 0.19 | 0.01 | 0.99 | – | – |
| 72 h | 400.68 | 104.99–703.52 | 0.69 ± 0.19 | 0.06 | 0.99 | 1030.30 | Synergistic |
| 96 h | 196.41 | 33.50–381.58 | 0.78 ± 0.21 | 1.09 | 0.78 | 691.34 | Synergistic |
| L. angustifolia + H. spicatum (24 h) | – | – | – | – | – | – | – |
| 48 h | 1700.38 | 905.35–9843.58 | 0.67 ± 0.19 | 0.84 | 0.84 | – | – |
| 72 h | 594.53 | 420.63–875.54 | 1.10 ± 0.20 | 1.00 | 0.80 | 875.43 | Synergistic |
| 96 h | 308.73 | 207.86–418.60 | 1.25 ± 0.21 | 2.70 | 0.44 | 607.70 | Synergistic |
| L. angustifolia + C. deodara (24 h) | – | – | – | – | – | – | – |
| 48 h | 1140.59 | 689.27–2961.81 | 0.73 ± 0.19 | 0.57 | 0.90 | – | – |
| 72 h | 585.95 | 302.74–1135.58 | 0.66 ± 0.19 | 0.34 | 0.95 | 888.25 | Synergistic |
| 96 h | 244.99 | 78.65–414.47 | 0.73 ± 0.19 | 0.89 | 0.83 | 765.81 | Synergistic |
| L. angustifolia + P. wallichiana (24 h) | 1322.93 | 837.00–2435.62 | 0.81 ± 0.19 | 0.15 | 0.98 | – | – |
| 48 h | 623.70 | 317.43–976.71 | 0.82 ± 0.19 | 0.07 | 0.99 | – | – |
| 72 h | 312.23 | 115.83–508.34 | 0.89 ± 0.20 | 0.06 | 0.99 | 1666.95 | Synergistic |
| 96 h | 214.19 | 103.57–320.54 | 1.41 ± 0.25 | 3.99 | 0.26 | 875.93 | Synergistic |
| L. angustifolia + J. communis (24 h) | 1376.68 | 899.68–2310.59 | 0.89 ± 0.19 | 0.42 | 0.94 | – | – |
| 48 h | 664.26 | 373.59–984.83 | 0.94 ± 0.20 | 0.32 | 0.96 | – | – |
| 72 h | 372.88 | 185.63–556.04 | 1.11 ± 0.21 | 1.43 | 0.70 | 1395.82 | Synergistic |
| 96 h | 251.79 | 126.34–371.48 | 1.43 ± 0.25 | 3.41 | 0.33 | 745.12 | Synergistic |
| Essential Oils | LC50 (µL/L) | Confidence Limits (µL/L) | Slope ± SE | Chi-Square | p-Value | Co – Toxicity Coefficient | Interaction Type |
|---|---|---|---|---|---|---|---|
| A. calamus (72 h) | 10,975.11 | 6875.74-27,344.73 | 1.10 ± 0.22 | 2.71 | 0.44 | – | – |
| 96 h | 1379.54 | 1001.07–1778.06 | 1.58 ± 0.22 | 3.03 | 0.39 | – | – |
| 120 h | 1158.42 | 863.34–1456.71 | 1.88 ± 0.25 | 3.85 | 0.28 | – | – |
| H. spicatum (72 h) | 10,448.32 | 7116.12–20,223.58 | 1.41 ± 0.24 | 1.69 | 0.64 | – | – |
| 96 h | 4875.82 | 3464.11–8042.55 | 1.14 ± 0.20 | 0.35 | 0.95 | – | – |
| 120 h | 1586.15 | 1099.21–2121.89 | 1.31 ± 0.21 | 2.28 | 0.52 | – | – |
| J. communis (72 h) | 9895.41 | 6865.13–18,255.56 | 1.45 ± 0.24 | 2.13 | 0.55 | – | – |
| 96 h | 5287.16 | 4018.85–7673.06 | 1.52 ± 0.22 | 3.19 | 0.36 | – | – |
| 120 h | 2312.36 | 1799.64–2954.15 | 1.63 ± 0.22 | 1.57 | 0.67 | – | – |
| J. recurva (96 h) | 5980.52 | 3945.11–12,352.29 | 0.97 ± 0.20 | 0.43 | 0.93 | – | – |
| 120 h | 1914.97 | 1413.88–2500.84 | 1.44 ± 0.21 | 2.35 | 0.50 | – | – |
| P. wallichiana (96 h) | 7918.09 | 5152.81–17,361.77 | 1.05 ± 0.21 | 1.30 | 0.73 | – | – |
| 120 h | 1694.87 | 1202.74–2250.79 | 1.35 ± 0.21 | 2.39 | 0.50 | – | – |
| C. deodara (72 h) | 4797.04 | 3485.81–7523.48 | 1.23 ± 0.21 | 0.04 | 0.99 | – | – |
| 96 h | 2598.47 | 1923.33–3536.37 | 1.30 ± 0.20 | 0.34 | 0.95 | – | – |
| 120 h | 1716.80 | 1243.41–2252.42 | 1.41 ± 0.21 | 1.62 | 0.65 | – | – |
| L. angustifolia (72 h) | 4316.34 | 3333.68–5798.88 | 1.54 ± 0.22 | 3.71 | 0.29 | – | – |
| 96 h | 1715.57 | 1346.35–2129.90 | 1.86 ± 0.23 | 4.65 | 0.20 | – | – |
| 120 h | 779.59 | 532.38–1009.98 | 1.95 ± 0.29 | 3.55 | 0.31 | – | – |
| A. calamus + L. angustifolia (24 h) | 396.54 | 256.96–618.49 | 0.92 ± 0.19 | 0.15 | 0.98 | – | – |
| 48 h | 201.22 | 114.02–292.26 | 1.02 ± 0.20 | 0.62 | 0.89 | – | – |
| 72 h | 141.89 | 86.94–195.48 | 1.39 ± 0.22 | 0.40 | 0.94 | 7734.94 | Synergistic |
| 96 h | 92.18 | 55.92–124.84 | 1.92 ± 0.31 | 3.04 | 0.39 | 1496.57 | Synergistic |
| A. calamus + C. deodara (24 h) | 509.92 | 330.43–795.31 | 0.92 ± 0.19 | 0.15 | 0.98 | – | – |
| 48 h | 258.76 | 146.62–375.82 | 1.02 ± 0.20 | 0.62 | 0.89 | – | – |
| 72 h | 182.46 | 111.79–251.37 | 1.39 ± 0.22 | 0.40 | 0.94 | 6015.08 | Synergistic |
| 96 h | 118.54 | 71.91–160.54 | 1.92 ± 0.31 | 3.04 | 0.39 | 1163.78 | Synergistic |
| A. calamus + H. spicatum (24 h) | 1196.11 | 828.88–1960.84 | 1.01 ± 0.20 | 0.09 | 0.99 | – | – |
| 48 h | 455.34 | 298.73–623.14 | 1.22 ± 0.21 | 1.28 | 0.73 | – | – |
| 72 h | 285.42 | 176.84–391.35 | 1.42 ± 0.23 | 1.38 | 0.71 | 3845.25 | Synergistic |
| 96 h | 201.90 | 128.37–269.01 | 1.94 ± 0.30 | 0.99 | 0.80 | 683.28 | Synergistic |
| A. calamus + J. communis (24 h) | 1545.98 | 1116.72–2442.67 | 1.19 ± 0.21 | 0.52 | 0.91 | – | – |
| 48 h | 911.85 | 648.64–1308.67 | 1.14 ± 0.20 | 0.03 | 0.99 | – | – |
| 72 h | 512.02 | 337.16–703.45 | 1.19 ± 0.20 | 0.27 | 0.97 | 2143.49 | Synergistic |
| 96 h | 275.64 | 160.70–386.58 | 1.37 ± 0.23 | 1.45 | 0.69 | 500.49 | Synergistic |
| A. calamus + J. recurva (24 h) | 2223.17 | 1513.69–4197.51 | 1.06 ± 0.20 | 0.11 | 0.99 | – | – |
| 48 h | 1238.06 | 860.39–1941.03 | 1.03 ± 0.20 | 0.08 | 0.99 | – | – |
| 72 h | 733.32 | 500.66–1020.75 | 1.14 ± 0.20 | 0.30 | 0.96 | 1496.63 | Synergistic |
| 96 h | 398.87 | 263.00–534.18 | 1.43 ± 0.22 | 3.41 | 0.33 | 345.86 | Synergistic |
| L. angustifolia + C. deodara (24 h) | 740.11 | 534.33–1099.13 | 1.15 ± 0.20 | 0.29 | 0.96 | – | – |
| 48 h | 432.83 | 300.38–595.76 | 1.19 ± 0.20 | 0.25 | 0.97 | – | – |
| 72 h | 279.06 | 189.12–372.06 | 1.39 ± 0.21 | 0.32 | 0.96 | 1546.74 | Synergistic |
| 96 h | 182.66 | 121.13–241.33 | 1.71 ± 0.25 | 3.03 | 0.39 | 939.21 | Synergistic |
| L. angustifolia + H. spicatum (24 h) | 1185.35 | 809.62–1954.79 | 0.97 ± 0.20 | 0.64 | 0.89 | – | – |
| 48 h | 649.55 | 401.60–966.17 | 0.95 ± 0.20 | 0.54 | 0.91 | – | – |
| 72 h | 354.18 | 159.61–550.13 | 0.89 ± 0.20 | 0.55 | 0.91 | 1218.69 | Synergistic |
| 96 h | 260.18 | 161.91–353.63 | 1.58 ± 0.25 | 4.39 | 0.22 | 659.38 | Synergistic |
| L. angustifolia + J. communis (24 h) | 1738.22 | 1188.13–3148.40 | 1.01 ± 0.20 | 1.31 | 0.74 | – | – |
| 48 h | 808.54 | 513.02–1235.52 | 0.92 ± 0.20 | 1.17 | 0.76 | – | – |
| 72 h | 449.20 | 271.46–633.03 | 1.13 ± 0.20 | 1.26 | 0.74 | 960.89 | Synergistic |
| 96 h | 284.89 | 183.15–381.61 | 1.65 ± 0.25 | 4.43 | 0.22 | 602.19 | Synergistic |
| L. angustifolia + J. recurva (24 h) | 2387.58 | 1602.20–4719.26 | 1.03 ± 0.20 | 1.65 | 0.65 | – | – |
| 48 h | 1089.42 | 741.91–1663.55 | 1.00 ± 0.20 | 1.03 | 0.79 | – | – |
| 72 h | 545.04 | 353.78–748.04 | 1.22 ± 0.21 | 1.85 | 0.60 | 791.93 | Synergistic |
| 96 h | 259.42 | 18.52–498.28 | 1.53 ± 0.25 | 5.61 | 0.13 | 661.31 | Synergistic |
| Essential Oils | RC50 (µL/L) | Confidence Limits (µL/L) | Slope ± SE | Chi-Square | p-Value |
|---|---|---|---|---|---|
| A. calamus (1 h) | 53.24 | 30.82–75.45 | 1.19 ± 0.16 | 3.03 | 0.39 |
| 2 h | 17.96 | 3.83–37.29 | 0.84 ± 0.16 | 1.64 | 0.65 |
| 3 h | 12.94 | 1.88–30.19 | 0.82 ± 0.17 | 1.63 | 0.65 |
| 4 h | 25.24 | 8.50–44.79 | 0.98 ± 0.17 | 1.95 | 0.58 |
| 5 h | 52.06 | 28.63–75.45 | 1.11 ± 0.16 | 4.50 | 0.21 |
| 24 h | 53.98 | 20.25–89.22 | 0.74 ± 0.14 | 4.85 | 0.18 |
| H. spicatum (1 h) | 208.36 | 168.43–254.45 | 1.35 ± 0.14 | 4.95 | 0.18 |
| 2 h | 109.26 | 84.43–134.10 | 1.50 ± 0.16 | 3.57 | 0.31 |
| 3 h | 126.80 | 93.78–160.43 | 1.21 ± 0.15 | 3.53 | 0.32 |
| 4 h | 198.92 | 158.10–245.46 | 1.27 ± 0.14 | 1.74 | 0.63 |
| 5 h | 254.71 | 205.33–316.63 | 1.26 ± 0.14 | 2.06 | 0.56 |
| 24 h | 293.77 | 248.19–350.26 | 1.64 ± 0.15 | 0.57 | 0.90 |
| J. recurva (1 h) | 176.79 | 140.60–216.57 | 1.33 ± 0.15 | 2.20 | 0.53 |
| 2 h | 219.77 | 183.66–261.37 | 1.59 ± 0.15 | 1.40 | 0.71 |
| 3 h | 238.35 | 205.39–276.16 | 1.95 ± 0.16 | 1.34 | 0.72 |
| 4 h | 342.45 | 293.15–404.90 | 1.82 ± 0.16 | 3.03 | 0.39 |
| 5 h | 359.55 | 304.70–431.45 | 1.69 ± 0.16 | 4.04 | 0.26 |
| 24 h | 309.75 | 243.99–402.39 | 1.11 ± 0.14 | 5.03 | 0.16 |
| P. wallichiana (1 h) | 221.50 | 188.69–258.94 | 1.80 ± 0.16 | 3.81 | 0.28 |
| 2 h | 269.48 | 234.55–310.31 | 2.09 ± 0.17 | 4.86 | 0.18 |
| 3 h | 255.00 | 215.86–301.41 | 1.68 ± 0.15 | 2.56 | 0.46 |
| 4 h | 381.69 | 332.91–442.18 | 2.17 ± 0.18 | 2.15 | 0.54 |
| 5 h | 507.48 | 416.70–645.97 | 1.50 ± 0.16 | 1.64 | 0.65 |
| 24 h | 955.15 | 686.08–1588.08 | 1.12 ± 0.15 | 1.02 | 0.79 |
| C. deodara (1 h) | 286.47 | 234.69–353.40 | 1.35 ± 0.15 | 1.37 | 0.71 |
| 2 h | 389.71 | 313.91–503.17 | 1.26 ± 0.15 | 0.82 | 0.84 |
| 3 h | 514.85 | 415.50–674.00 | 1.37 ± 0.15 | 1.13 | 0.77 |
| 4 h | 331.02 | 276.27–402.95 | 1.51 ± 0.15 | 0.53 | 0.91 |
| 5 h | 496.27 | 398.37–653.91 | 1.31 ± 0.15 | 0.45 | 0.93 |
| 24 h | 425.75 | 335.03–572.70 | 1.13 ± 0.14 | 5.10 | 0.16 |
| L. augustifolia (1 h) | 174.41 | 147.28–203.85 | 1.82 ± 0.16 | 5.08 | 0.17 |
| 2 h | 187.60 | 151.89–227.18 | 1.42 ± 0.15 | 3.99 | 0.26 |
| 3 h | 228.58 | 187.98–276.11 | 1.44 ± 0.15 | 3.33 | 0.34 |
| 4 h | 408.32 | 347.72–489.11 | 1.80 ± 0.16 | 0.37 | 0.95 |
| 5 h | 407.11 | 328.26–526.27 | 1.27 ± 0.15 | 2.19 | 0.53 |
| 24 h | 735.70 | 593.16–977.46 | 1.60 ± 0.17 | 2.78 | 0.43 |
| J. communis (1 h) | 213.21 | 144.14–302.11 | 0.75 ± 0.14 | 1.03 | 0.79 |
| 2 h | 350.99 | 266.29–489.15 | 0.95 ± 0.14 | 3.70 | 0.30 |
| 3 h | 305.05 | 225.62–428.66 | 0.87 ± 0.14 | 0.18 | 0.98 |
| 4 h | 411.56 | 312.15–586.36 | 0.96 ± 0.14 | 0.46 | 0.92 |
| 5 h | 356.47 | 253.87–548.20 | 0.76 ± 0.14 | 2.30 | 0.51 |
| 24 h | 736.82 | 512.04–1327.91 | 0.87 ± 0.14 | 5.05 | 0.17 |
| Essential Oils | RC50 (µL/L) | Confidence Limits (µL/L) | Slope ± SE | Chi-Square | p-Value |
|---|---|---|---|---|---|
| A. calamus (1 h) | 26.04 | 7.99–47.60 | 0.89 ± 0.16 | 0.15 | 0.99 |
| 2 h | 24.69 | 8.79–43.03 | 1.05 ± 0.18 | 0.45 | 0.93 |
| 3 h | 12.94 | 1.88–30.19 | 0.82 ± 0.17 | 1.63 | 0.65 |
| 4 h | 10.08 | 0.25–56.83 | 0.44 ± 0.14 | 1.74 | 0.63 |
| 5 h | 21.67 | 0.16–64.21 | 0.40 ± 0.14 | 0.79 | 0.85 |
| 24 h | 118.91 | 36.91–205.00 | 0.48 ± 0.13 | 0.28 | 0.96 |
| H. spicatum (1 h) | 30.60 | 8.86–56.30 | 0.79 ± 0.15 | 0.10 | 0.99 |
| 2 h | 32.88 | 7.89–63.02 | 0.69 ± 0.14 | 0.48 | 0.92 |
| 3 h | 44.39 | 7.76–87.66 | 0.55 ± 0.14 | 0.32 | 0.96 |
| 4 h | 98.06 | 25.75–172.36 | 0.48 ± 0.13 | 1.25 | 0.74 |
| 5 h | 86.14 | 5.75–176.71 | 0.35 ± 0.13 | 1.85 | 0.60 |
| 24 h | 226.85 | 151.57–329.13 | 0.72 ± 0.14 | 4.57 | 0.21 |
| J. recurva (1 h) | 163.57 | 124.14–206.15 | 1.16 ± 0.14 | 1.36 | 0.71 |
| 2 h | 216.24 | 174.29–265.40 | 1.31 ± 0.15 | 1.80 | 0.61 |
| 3 h | 320.54 | 264.62–395.06 | 1.41 ± 0.15 | 3.29 | 0.35 |
| 4 h | 393.61 | 332.45–476.02 | 1.68 ± 0.16 | 3.91 | 0.27 |
| 5 h | 438.79 | 366.75–540.90 | 1.60 ± 0.16 | 0.30 | 0.96 |
| 24 h | 383.51 | 313.39–484.14 | 1.35 ± 0.15 | 3.72 | 0.29 |
| P. wallichiana (1 h) | 225.56 | 181.99–277.35 | 1.30 ± 0.14 | 0.08 | 0.99 |
| 2 h | 262.18 | 207.52–332.63 | 1.15 ± 0.14 | 3.79 | 0.28 |
| 3 h | 363.70 | 291.20–470.43 | 1.20 ± 0.14 | 1.87 | 0.60 |
| 4 h | 432.24 | 326.73–623.50 | 0.96 ± 0.14 | 2.41 | 0.49 |
| 5 h | 481.32 | 369.81–681.46 | 1.06 ± 0.14 | 1.16 | 0.76 |
| 24 h | 947.27 | 644.05–1852.50 | 0.90 ± 0.14 | 0.59 | 0.90 |
| C. deodara (1 h) | 610.61 | 452.44–941.18 | 1.00 ± 0.14 | 4.75 | 0.19 |
| 2 h | 553.77 | 414.55–832.24 | 0.99 ± 0.14 | 0.34 | 0.95 |
| 3 h | 629.00 | 487.84–888.89 | 1.23 ± 0.15 | 2.01 | 0.57 |
| 4 h | 463.66 | 364.53–629.24 | 1.16 ± 0.14 | 3.48 | 0.32 |
| 5 h | 600.86 | 473.37–824.52 | 1.28 ± 0.15 | 3.30 | 0.35 |
| 24 h | 628.88 | 443.26–1087.98 | 0.84 ± 0.14 | 4.19 | 0.24 |
| L. augustifolia (1 h) | 113.67 | 88.26–139.13 | 1.50 ± 0.16 | 4.86 | 0.18 |
| 2 h | 176.35 | 147.17–208.02 | 1.69 ± 0.16 | 5.20 | 0.16 |
| 3 h | 228.58 | 187.98–276.11 | 1.44 ± 0.15 | 3.33 | 0.34 |
| 4 h | 333.52 | 264.64–433.91 | 1.14 ± 0.14 | 5.18 | 0.16 |
| 5 h | 392.81 | 306.70–531.41 | 1.08 ± 0.14 | 0.68 | 0.88 |
| 24 h | 681.78 | 558.79–879.86 | 1.69 ± 0.17 | 2.19 | 0.53 |
| J. communis (1 h) | 118.35 | 79.08–158.04 | 0.98 ± 0.14 | 2.07 | 0.56 |
| 2 h | 243.59 | 182.03–324.53 | 0.94 ± 0.14 | 0.27 | 0.97 |
| 3 h | 334.26 | 253.74–461.23 | 0.95 ± 0.14 | 0.24 | 0.97 |
| 4 h | 398.11 | 307.99–547.21 | 1.04 ± 0.14 | 0.63 | 0.89 |
| 5 h | 242.53 | 167.66–347.66 | 0.76 ± 0.14 | 3.43 | 0.33 |
| 24 h | 617.11 | 428.88–1104.77 | 0.80 ± 0.14 | 4.75 | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, H.; Deeksha; Urvashi; Reddy, S.G.E. Insecticidal and Detoxification Enzyme Inhibition Activities of Essential Oils for the Control of Pulse Beetle, Callosobruchus maculatus (F.) and Callosobruchus chinensis (L.) (Coleoptera: Bruchidae). Molecules 2023, 28, 492. https://doi.org/10.3390/molecules28020492
Gupta H, Deeksha, Urvashi, Reddy SGE. Insecticidal and Detoxification Enzyme Inhibition Activities of Essential Oils for the Control of Pulse Beetle, Callosobruchus maculatus (F.) and Callosobruchus chinensis (L.) (Coleoptera: Bruchidae). Molecules. 2023; 28(2):492. https://doi.org/10.3390/molecules28020492
Chicago/Turabian StyleGupta, Himanshi, Deeksha, Urvashi, and S. G. Eswara Reddy. 2023. "Insecticidal and Detoxification Enzyme Inhibition Activities of Essential Oils for the Control of Pulse Beetle, Callosobruchus maculatus (F.) and Callosobruchus chinensis (L.) (Coleoptera: Bruchidae)" Molecules 28, no. 2: 492. https://doi.org/10.3390/molecules28020492