Pipa malá (Pipa parva)
Pipa parva Ruthven and Gaige, 1923 - Sabana Surinam Toad

Young adult male
Forein language names: Kleine Wabenkröte (German), pipa malá (Czech), pipa pequeňa (Spanish), Dvaergpipatudse (Dutch)
Charakteristic features
It is fully aquatic species. It is a small species within the genus Pipa and a medium-sized within the all family Pipidae. Body longht (snout-vent) is usually from 35 to 50 mm. Females reach a larger adult size than males. The body is dorsoventrally flattened. Females generally have plumper body with wider hips. The front limbs are significantly weaker than the hind limbs. Front limbs are without webs, hind limbs are with webs between all toes and along the entire their length. There are always four asymmetrically located projections on the tips of the fingers of the front limbs. The small head smoothly connects to the body with minimal possibility of rotation. Eyes are small, inexpressive, directed upwards with a round pupil, without eyelids. Snout slightly blunt in dorsal view. The skin is slightly warty on dorsal side. Coloration is bland, uniform, almost identical in both sexes, variability relatively small and mostly on the belly. The entire upper side of the body, head and limbs are colored a uniform dark gray with darker small spots. The underside of the body is uniformly dirty white with small gray spots, especially on the rump and chest.
Taxonomy
The species was described in February 1923 as the third species known at that time, after the Pipa pipa described as early as 1758 (Linnaeus) and the Pipa snethlageae described in 1914 (Müller). The fact that this is a new species of pipa was not difficult to recognize at first glance by the size and overall character of the body. The complete scientific article description can be found here. Scientific description of the species was published by herpetologist and president of University of Michigan Alexander Grant Ruthven and herpetologist Helen Beulah Thompson Gaige from Museum of Zoology, University of Michigan. The description was made on base of collections from nature dated 5th May (possibly 28th April) in 1922 by M. A. Carriker Jr. in the village Sabana de Mendoza (Trujillo State) on the eastern shore of Lake Maracaibo in Venezuela (covered drainage channel). However, the exact location of the collection point is not known. The description of the species was published in the journal "Ocassional papers of the museum of zoology" of the University of Michigan. The description is quite brief and was made on the basis of dead individuals fixed in alcohol.
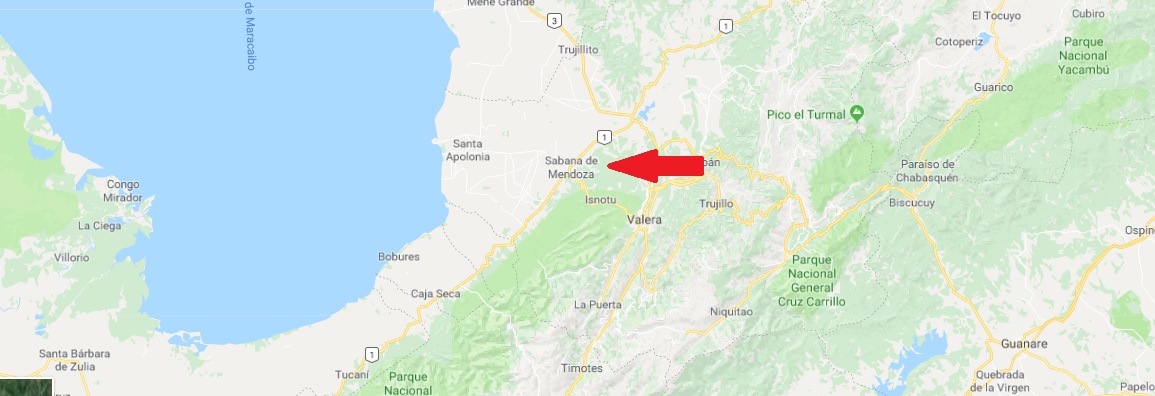
Village or small town Sabana de Mendoza in Venezuela (red arrow), where is type locality od Pipa parva.
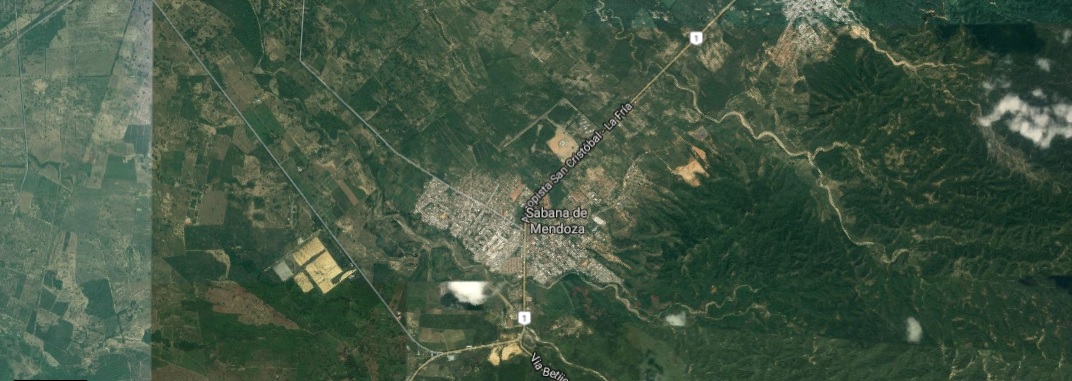
Bližší satelitní pohled na město Sabana de Mendoza, v jehož blízkosti se nalézá typová lokalita pipy malé (P. parva).
Currently, the holotype of the species is stored in the Zoological Museum of the University of Michigan under the number 57443. In addition to the holotype, there are 6 other paratypes (paratotypes) of the type series. From these six, one paratype (paratotype) is missing and one is deposited at Harvard University in an unspecified exchange.

 Holotype of Pipa parva (female) in dorsal (left) and ventral views deposited at the University of Michigan, USA (source University of Michigan, https://quod.lib.umich.edu).
Holotype of Pipa parva (female) in dorsal (left) and ventral views deposited at the University of Michigan, USA (source University of Michigan, https://quod.lib.umich.edu).
Samotný latinský druhový název parva znamená v překladu z latiny "malý" (parvus). Svojí velikostí se řadí mezi tzv. malé pipy (mikropipy) společně s druhem pipou Myersovou (P. myersi), která byla svojí podobností považována celých 61 let za pipu malou, než byla vyčleněna Lindou Truebovou z Kansaské univerzity jako samostatný druh (panamské a kolumbijské populace). Pipa Myersova je tak sesterský, pravděpodobně blízce příbuzný druh pipy malé. Rozdíly obou druhů jsou velmi malé a ustanovení druhu pipy Myersovy bude vyžadovat další revizi, zejména genetického charakteru, zda-li je vyčlenění tohoto druhu oprávněné a nejedná se například pouze o pohttps://quod.lib.umich.eduruh pipy malé.
Pipa malá nevytváří žádné poddruhy, respektive nebyly doposud vědecky popsány.
Poznámka k historii rodového označení. V roce 1925 Noble ustanovil nový rod Protopipa (vedle do té doby jediného známého rodu Pipa) s typovým druhem pipou venezuelskou (Pipa aspera). Do tohoto rodu navrhl zařadit kromě tohoto druhu i pipu malou. Do rodu Protopipa tak navrhoval zařadit druhy, které nemají celkový charakter těla zploštělý a nemají kožní výběžky. V rodu Pipa navrhl zachovat druhy pipu velkou (Pipa pipa), pipu drobnou (Pipa snethlagea) a Pipa pernigra (v současné době neuznávaný druh, synonimum Pipa pipa). Momentálně není rod Protopipa uznáván, všechny druhy pip náleží pouze do rodu Pipa. Celý článek zde.
Genetická data
Karyotyp obsahuje v diploidní sadě (2n) 30 chromozomů.
Stavba těla a rozměry
Svojí velikostí se jedná o malý druh pipy a spolu s pipou myersovou patří mezi nejmenší pipy vůbec. Literatura uvádí, že samci i samice dosahují délky těla od 27 do 44 mm (měřeno od špičky čenichu po střed kloaky). Dle jedinců odchovaných v zajetí může být délka těla i o několik milimetrů větší, konkrétně u samic 60 mm a u samců 53 mm (vlastní měření).
Přední končetiny jsou výrazně slabší a kratší oproti končetinám zadním. Po celé délce jsou přibližně stejně široké a směřuji dopředu paralelně s tělem a hlavou. U samců jsou přední končetiny, zejména předloktí, přibližně o třetinu silnější oproti samicím. Čtyři prsty jsou přibližně stejně dlouhé a stejně široké, rovnoměrně symetricky kuželovitě se zužující. Klouby nejsou na prstech patrné. Prsty jsou v poslední třetině volně pasivně (vlivem okolních předmětů) ohebné do všech stran. Zvířata nemohou prsty aktivně ohýbat nebo jen velmi omezeně. Dovedou prsty pouze vůči sobě navzájem rozvírat a přitisknout, případně mírně všechny společně proximálně ohýbat směrem k ústům (přihrnování potravy). Dospělí a zejména starší jedinci mohou mít v klidu přirozeně poslední třetinu délky prstů distálně ohnutou. Mezi prsty nejsou vyvinuté plovací blány, u báze prstů ale je jejich nezřetelný náznak přítomen. Špičky prstů jsou ukončeny charakteristickým rozvětvením. Na každém prstu je čtveřice výběžků. Ty jsou asymetricky rozloženy na špičkách prstů. Všechny výběžky jsou přibližně stejné velikosti. Dva vnější výběžky jsou umístěny na špičce prstu a směřují distálně dopředu. Zbylé dva vnitřní výběžky nejsou úplně na špišce prstu a směřují mírně proximálně. Výběžky konce prstů jsou na jedincích přítomny už od metamorfózy. Charakter konce prstů je pro tento druh specifický a výlučný a dle literatury tento znak není u žádného jiného druhu rodu Pipa. Jedná se tak o základní diagnostický znak, podle kterého pipu malou odlišíme od všech ostatních druhů, zejména od vzhledově příbuzné pipy brazilské (Pipa carvalhoi), která má všechny čtyři výběžky umístěny symetricky na špičkách prstů a sesterského druhu pipy Myersovi (Pipa myersi), která má pouze tři prstové výběžky. Předpokládá se, že výběžky mají četná nervová zakončení a pomáhají jedinci lokalizovat hmatově potravu v často kalných vodách.Samci nedisponují na vnitřní straně předních končetinách žádnými viditelnými pářícími mozoly. Tento znak tak nelze použít pro rozlišení pohlaví.

Finger detail of left hand of adult male with charakteristic prostrusions on finger tips.
Zadní končetiny nedisponují žádným vitřním ani vnějším patním (metatarsálním) hrbolem. Stejně tak nedisponují žádnými brbolky na spodní straně prstů a chodidla.
Oči jsou relativně malé symetricky kulaté o průměru cca 1,5 mm a směřují nahoru. Zřítelnice je černá, mírně oválná. Ve vrcholové části neostře lomená, ve spodní části s drobným výběžkem směrem dolů, který narušuje oválnou symetrii. Duhovka je po celém obvodu stejně široká. Šířka duhovky je přibližně jako poloměr zřítelnice v nejužším místě. Duhovka je špinavě zlatavá a jemně granulovaná. Vzdálenost mezi okem a nejbližším okrajem nozdry je přibližně 2,5 násobek průměru oka (zřítelnice + duhovka).
Nozdry jsou štěrbinovité umístěné na špičce čenicha.

Dospělý samec dýchající u hladiny.
Druh disponuje několika řadami laterálních orgánů. Jeden pár se táhne od čenichu k přednímu okraji každého oka. Přidružuje se k němu jeden dodatečný nebo dva spojené orgány u dorsálního nebo ventrálního okraje oka. Další řady s vertikálním uložením orgánů začínají po stranách hlavy u čelistního klubu a táhnou směrem dozadu přes předloktí a podél ventrální strany slabin. Párová podélná řada se táhne od spánkové oblasti k hřbetu. Další podélná řada se táhne od každého předloktí podél hřbetní strany boku.
Zbarvení
Základní zbarvením dorsální strany je tmavě šedočerné, šedé až tmavě hnědé. Na tomto podkladu jsou nevýrazné tmavší skvrnky s nepravdielnými okraji o velikosti několka milimetrů v průměru. Dorsální zbarvení hlavy, trupu i končetin je shodné, mezi těmito partiemi těla nejsou rozdíly. Rozdíly ve zbarvení mezi pohlavými jsou minimální. Variabilita ve zbarvení dorsální strany těla je mezi jedinci minimální. Celkově je zbarvení svým charakterem kryptické, má za úkol jedince maskovat při pohledu shora. Tmavé dorsální zbarvení přechází na bocích těla i končetin náhle do světlého zbarvení ventrální strany. Přechod není doprovázen žádným lineárním prvkem.


Dorsální pohled na dospělou samici (vlevo) a dospělého samce (jedinci nejsou zobrazeni ve vzájemném velikostním poměru).
Celá ventrální strana těla (hlava, trup, končetiny) má základní podkladní barvu plošně světle špinavě bílou. Na tomto podkladě jsou roztroušené šedivé skvrnky o velikosti obvykle do několika málo milimetrů. Hustota skvrnek obvykle roste směrem od zadní části k hlavě. Mezi jedinci je velká variabilita v odstínu a intentitě šedé barvy, počtu, velikosti a rozložení těchto šedých skvrnek.

Zbarvení ventrální strany těla dospělé samice.
Pohlavní dvojtvárnostPohlavní rozdíly, dvojtvárnost (sexuální dimorfismus) je relativně malé. Pro laika jsou obě pohlaví na první pohled relativně stejná nebo jen málo rozdílné. Dospělé samice jsou větší a zavalitější oproti samcům, kteří jsou pohledově gracilnější, štíhlejší a menší. Dospělí samci mají přibližně o třetinu silnější přední končetiny v celé své délce. Samci nedisponují jakýmikoli pářícími mozoly na předních končetinách, u obou pohlaví tak z vnitřní strany končetin nenajdeme žádné barevné nebo strukturální rozdíly. Ve zbarvení těla není též žádný rozdíl mezi oběma pohlavími. Samice mají větší kloaku a větší bradavice kůže na horní straně těla. O strukturálním rozdílu v pokožce samic a samců pojednává práce Kretschmer & Greven 2009.


Detail kloaky u dospělé samice (vlevo) a dospělého samce. Klokaka je u samic zřetelně větší.
Hlasové projevy, vokalizace
Hlasově se ozývají výlučně samci a to pod vodou. Smysl vokalizace je přilákat samice k páření. Samci nemají žádné rezonanční měchýřky, tedy se u nich pohledově vokalizace nijak neprojevuje. Hlas je vytvářen speciálním párem kůstek v hrdle, kterými jedinec o sebe navzájem "ťuká". Jejich hlas je relativně tiché trilkování nebo vrzání. Ukázka:

Geographical distribution
Distribution of Pipa parva is mainly in Venezuela. The species occupies also a small part of eastern Colombia. In Venezuela it occurs naturally in the northwestern part in the states of Zulia. This state covers majority of whole species distribution especially on west, north and east. The species is also in state Trujillo (eastern part of the range), Tachira (southern part of the range, Barroso & Fuentes 2000), Mérida (southern part of the range, Heatwole 1963) and probably also in Falcón state (northeastern part of the range). In the west, its occurrence crosses the border with Colombia (Lynch 2006), where it occurs in the extreme east in the state of Santander (Department Norte de Santander). But the main occurrence is in Venezuela around or close to lake Maracaibo (Maracaibo depression or basin) sandwiched between Serranía del Perijána (Perija range) in the west and Cordillera de Mérida in the south and east (both belong to the Eastern Cordillera). The mountain range probably forms a natural geographical barrier for spreading of the species to other adjacent territories. Occurrence is mostly hypsometrically limited to lower altitudes from 0 to 300 m above sea level (Maracaibo basin). Nevertheless, it can also occur at higher altitudes on the edge of its range, as evidenced by, for example, the most southerly documented occurrence in the vicinity of the city of La Grita in the state of Tachira with a reported altitude of 1440 m above sea level (Barroso & Fuentes 2000).
V rámci rodu Pipa se jedná pravděpodobně o druhý nejmenší areál výskytu. Ještě menší má pravděpodobně pouze pipa Myersová (Pipa myersi) z Panamy a snad i z Kolumbie, i když zde je její výskyt více než pochybný. Původně byly panamské a kolumbijské populace považovány za pipu malou, v roce 1984 ale byly tyto populace vyčleněny jako samostatný druh pipa Myersova (Pipa myersi).
Kromě přirozeného výskytu byla pipa malá introdukovaná do západní oblasti jezera Valencia ve Venezuelském státě Carabobo. Výskyt je zde pravděpodobně způsoben antropogenním vlivem s úmyslným vysazením pipa malé za účelem jejího samovolného množení s následným odlovem a prodejem zvířat pro akvaristický trh, případně jako neúmyslný výsadek s následným nekontrolovatelným, šířením druhu. Výskyt na tomto území je doložen poprvé v roce 1996 (Royero & Hernandéz, 1996). Panuje zde obava z dalšího šíření druhu zejména do povodí Orinoka. Tato skutečnost potvrzuje, že úmyslná i neúmyslná introdukce jakéhokoli organismu na jakékoli jiné území mimo jeho původní výskyt je nepřijatelné a nežádoucí!
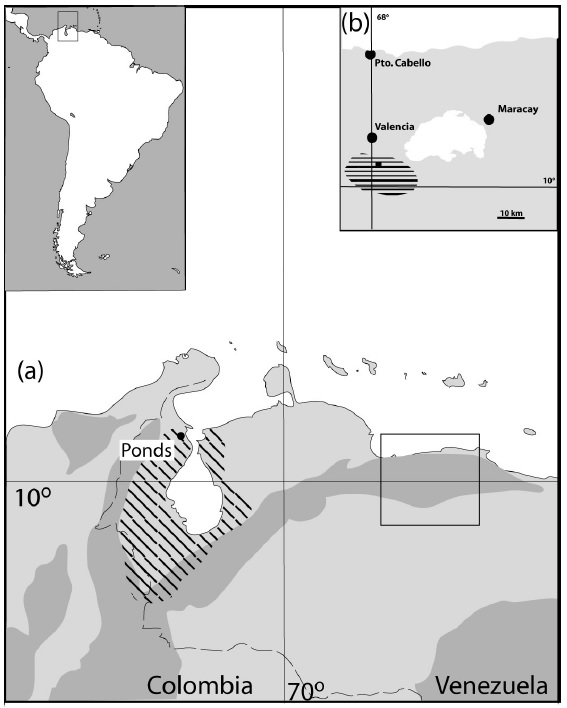
Přirozený výskyt pipy malé v okolí jezera Maracaibo dle Measey & Royero, 2005 - šikmo šrafovaně a introdukce v západní části jezera Valencia - vodorovně šrafovaně.
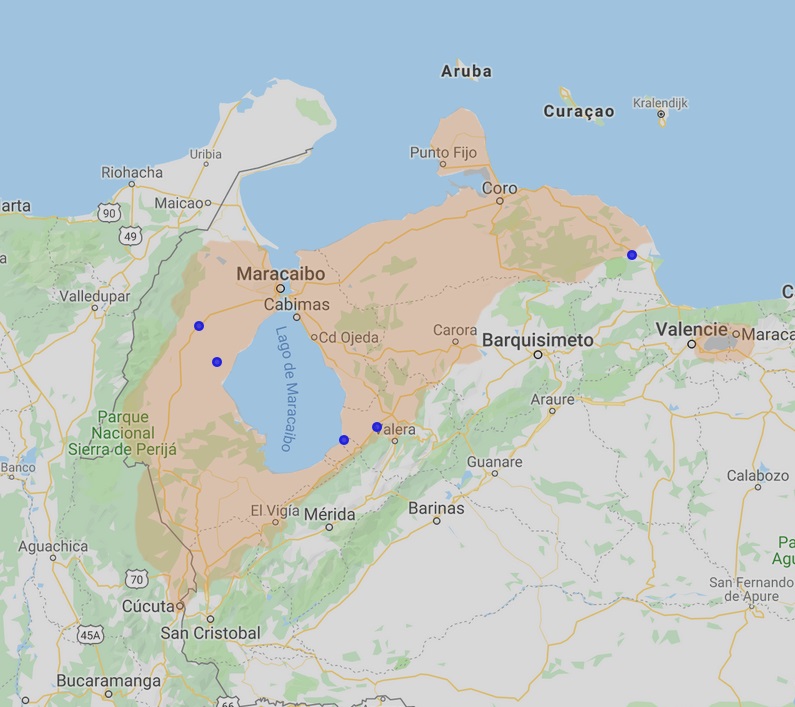
Další dostupná mapa výskytu pipy malé dle AmhibiaWeb, 2019. Oproti výskytu dle Measey & Royero, 2005 je zde uváděn daleko větší rozšíření, které zahrnuje i celou nížinu na východ od jezera Maracaibo (srovnej s předchozí mapou). Přirozený areál výskytu by tak byl téměř dvojnásobný a zahrnoval by i venezuelské státy Falcón a Lara. Výskyt pipy malé i ve východní části dokládá nález z roku 1942 (modrá tečka zcela vpravo). Zda se jedná ve východní části o historický výskyt či jde spíše o nedostatek faunstických dat a pozorování druhu vyžaduje další terénní průzkum.
Syntopicky se s pipou malou v přirozených oblastech rozšíření pravděpodobně nevyskytuje žádný jiný druh rodu Pipa. Nejblíže k tomu má pipa velká (P. pipa), jejíž areál zasahuje od jihu do západní Venezuely a od výskytu s pipou malou jí dělí přibližně 50 km (AmphibiaWeb, 2019). Syntopický výskyt tak není vyloučen.
Biotop
Pipa malá osidluje stojaté vodní plochy jako jezera, rybníky, tůně, vodní příkopy a podobně. V tekoucích vodách se pravděpodobně nevyskytuje nebo jen omezeně (výskyt v tekoucích vodách není nikde v literatuře zmíněn). Obývá jak přirozená vodní tělesa, tak antropogenní vodní plochy. Krajina s výskytem je otevřená, převážně travnatá. Výjimečně opouští vodu, pravděpodobně při mimořádně nepříznivých životních podmínkách. Tento fakt pravděpodobně přispívá k rychlému šíření druhu v okolí jezera Valencia, kde byl druh vysazen a kde je považován za škůdce. Rychle se zde množí a představuje konkurenci chovaným rybám.
V přírodě je pravděpodobně poměrně obtížně pozorovatelná díky své plachosti a noční aktivitě. Ráda se ukrývá v sedimentech dna nebo vodních rostlinách.
Teplota i 32,9 °C, pH 10,54 (Measey & Royero, 2005)
Potrava
Jedinou prací zabývající se studiem kvality a kvantity potravy pipy malé v přírodě je Measley & Royero, 2005. U 42 jedinců z invazivní populace v okolí jezera Valencia ve venezuelském státě Carabobo. Valnou část potravy tvořily larvy pakomárů (Chironomidae). Ty se vyskytovaly u 93% vzorků. Následovaly larvy jepic (Ephemeroptera) a brouků (Coleoptera) s výskytem u 33% vzorků u obou skupin, byly zjištěny dva typy korýši lasturnatek (Ostracoda) s výskytem u 21 a 24% vzorků. Též byla zjištěna vajíčka vodních šneků u 2% vzorků. U pěti jedinců z přirozeného výskytu z okolí jezera Maracaibo byly zjištěny pouze vodní šneci okružáci (Planorbidae) s výskytem u 40% vzorků. Z výsledku je zřejmé, že pipa malá je potravní generalista. Složení potravy je zřejmě dáno aktuální biotopovou nabídkou.
Chování
Jde o klidný a neagresivní druh s převážně noční aktivitou. Přes den se zvířata převážně ukrývají a odpočívají na dně mezi kořeny, listím a rostlinami a nevykazují přílišnou aktivitu. Vidět jsou zejména když se plavou nadechnout k hladině. Vyhřívání u hladiny prakticky neprovádí. Pokud jim to podmínky dovolí, mohou být v natažené vertikální poloze opírající se zadními končetinami o dno nebo předměty v nádrži a s dozdrami nad hladinou. V noci pak aktivně plavou a hledají a přijímají potravu. Rozmnožování též probíhá v průběhu noci, výjimečně i v ranních hodinách za světla. Zvířata nejsou vůči sobě ani jiným zvířatům agresivní. V průběhu roku se nevyskytuje zimování či estivace. Jedinci se páří celoročně. Samci vykazují neagresivní teritoriální chování vůči jiným samcům svého druhu, které se projevuje vzájemným přetlačováním samců.
Význam a ochrana
Druh je potenciálně ohrožen ztrátou a změnami biotopů svého výskytu. Riziko vlivu se zdá být ale malé. Druh se tak jeví početně jako stabilní a perspektivní. Pipa malá je zdá se poměrně plastický druh, který osídluje i polopřirozené až umělé biotopy a můžeme u ní hovořit o probíhajícím procesu synantropizace.
Rozmnožování

Pár v amplexu
Obě pohlaví dosahují pohlavní dospělosti přibližně ve věku tří měsíců (vlastní pozorování u jedinců odchovaných v zajetí při teplotě 26 °C a každodenním krmení). To je poměrně brzy a výrazně k tomu přispívá malá velikost v dospělosti. Samci se ozývají přerušovaným cvrčivým nebo vrzavým hlasem. Ten není nijak hlasitý slyšitelný maximálně na několik málo desítek metrů. Samci uchopují samice před zadními končetinami (amplexus inguinalis). Samice jsou značně k páření odmítavé a snaží se samce trhavými pohyby těla a napřímením zadních končetin zbavit. Po cca 12 - 24 hodinách od utvoření amplexu zduří samici na zádech kůže, což je základní předpoklad pro pozdější zanoření vajíček do kůže. Současně samici zduří kloaka. Velikost vajíček je přibližně 1,5 mm. Jsou světle jednolitě krémově bílá. Rosolovitý obal vlastního vajíčka je redukován na minimum. Pulci dosahují celkové délky až 40 mm. Plavou převážně ve vodním sloupci s hlavou dolů a drobným kmitáním konce ocásku. Mají plně pod kontrolou a ovládají hydrostatický vztlak, tedy vertikální pohyb vodním sloupcem. K tomu používají plíce a ocásek. Potravu získávají filtrováním vody pomocí hyobranchiálního pumpování - nasávají vodu ústním otvorem a vypuzují za hlavou, přičemž z vody filtrují potravu. Pokud je ve vodním sloupci nedostatek potravy, snaží se vířit dno, aby se jim přísun potravy zvýšil.
Video pulců ve stáří 12 hodin od "narození":

Populační data
Nejsou známy.
Chov v zajetí

Ukázka biotopové chovné nádrže s dospělou samicí v popředí.
Chov v zajetí je velmi nenáročný, jelikož tento druh nemá nějaké specifické či vyhrněné nároky na prostředí. Je vhodný i pro úplné začátečníky. Chováme jej výlučně ve vodě v akváriích či jiných vhodných nádržích bez nutnosti souše, na kterou až na ojedinělé výjimky nevylézá. Na jednu žábu počítáme minimálně s 5 litry vody. Jakožto tropický druh vyžaduje celoročně teplotu vody ideálně v rozsahu 23 - 27 °C. Druh nesnáší teploty pod 20 °C. Teploty pod cca 15°C jsou život ohrožující. Topná tělíska vybavíme kryty, aby se o ně jedinci nepopálili, když se za nimi snaží ukrýt - to platí zejména při chovu mladých jedinců. Ideální je mít výšku vodního sloupce 10-15 cm, může být ale i vyšší. Jde o druh s noční aktivitou, který se přes den ukrývá, proto nádrž vybavíme úkryty, ideálně kořeny, případně bukovým a dubovým listím, do kterých se jedinci rádi ukrývají pro pocit bezpečí (není podmínkou). Žáby ale můžeme chovat i v klasicky vybavenm akváriu. Filtrace není bezpodmnečně nutná, žáby snáší i poměrně silné organické znečištění vody. Dno nádrže ůže být se substrátem (písek, neostré kamínky), může být ale i holé bez jakéhokoli substrátu. Ideální je vybavit nádrž vznášejícími se a plovoucími rostlinami (vodní mor kanadský, stolístek, okřehek, voďankovec apod.). Rostliny tlumí intenzitu akvarijního osvětlení a poskytují jedincům další úkryty, kde se cítí v bezpečí. Výměna vody a čistění chovné nádrže nemusí být nijak intenzivní, záleží na konkrétních podmínkách chovu (intenzitě krmení, počtu zvířat) a dle aktuální situace obvykle jednou za týden až měsíc vyměníme třetinu až polovinu objemu nádrže. Pipu malou krmíme převážně bezobratlými živočichy jako jsou zejména patentky, koretry a černé komáří larvy. \možnou potravou mohou být i blešivci (rod Gammarus), veléi dafnmi (živé i mražené), žížaly. Výjimečně a omezeně můžeme podávat i rybí maso, kuřecí játra a kuřecí srdce. Touto potravou ale nedoporučuji krmit zvířata trvale, protože se nejdená o přirozenou potravu. Mladé jedince po metamorfóze krmíme dvakrát denně, dospělce jednou za jeden až dva dny. Doporučuji chovat obě pohlaví odděleně. Důvodem je trvalá snaha samců pářit se se samicemi, které jsou v důsledku toho velmi plaché a ve stresu. Nedoporučuji chovat tento druh s rybami..
Podobné druhy
Když pomineme "sesterský" druh pipu Myersovu (Pipa myersi), která se v zajetí prakticky nechová, tak je pipa malá vzhledově i biologií velmi podobná pipě brazilské (Pipa carvalhoi). Zásadní vzájemné morfologické rozdíly obou druhů:
- p. brazilská je v dospělosti větší oproti p. malé, to platí pro obě pohlaví
- u p. brazilské není tak výrazný mezipohlavní rozdíl ve velikosti oproti p. malé
- p. brazislká má viditelnou mezioční (interorbitální) kožní lištu, p. malá ji postrádá
- p. brazilská má vizuálně výraznější postranní orgán oproti pipě malé, zejména na horní straně boků
- p. brazilská má tenší a delší přední končetiny oproti p. malé, to platí zejména pro samce
- p. brazilská má konce prstů předních končetin symetricky zakončeny čtyřmi výběžky. Pipa malá má též na každém prstu předních končetin čtyři výběžky, ty jsou ale asymetricky umístěny (blíže viz popis výše).
Oby druhy mají alopatrický výskyt, rozdílné zvukové projevy, mírně odlišné zbarvení, chování atd.
Použitá a doporučená literatura
Dunn, E.R. 1948. American frogs of the family Pipidae. American Museum Novitates 1384: 1–13.
Gines, H. 1958. Representantes de la familia Pipidae (Amphibia: Salientia) en Venezuela. Mem. Soc. Cien. Natur. La Salle, 18 (1): 5-18.
Heatwole, H. 1963. The frog genus Pipa in Panama. Copeia 1963: 436 - 438.
Moravec, J. 2001. České názvy živočichů IV., Obojživelníci (Amphibia), Národní Muzeum, Praha.